Neurophysiologie du comportement et cognition : intérêt de l’approche comparative
- Par Hugo Cousillas
- et Pierre Deleporte
Pages 131 à 143
Citer cet article
- COUSILLAS, Hugo
- et DELEPORTE, Pierre,
- Cousillas, Hugo.
- et al.
- Cousillas, H.
- et Deleporte, P.
https://doi.org/10.3917/biosy.027.0131
Citer cet article
- Cousillas, H.
- et Deleporte, P.
- Cousillas, Hugo.
- et al.
- COUSILLAS, Hugo
- et DELEPORTE, Pierre,
https://doi.org/10.3917/biosy.027.0131
Introduction
1 La biologie se donne pour but de comprendre la structure et le fonctionnement des êtres vivants, avec un intérêt privilégié bien compréhensible pour l’espèce humaine. L’un des défis les plus grands pour les biologistes est de comprendre le comportement humain, et le fonctionnement même de « l’esprit » humain, c’est-à-dire les capacités cérébrales de l’espèce la plus complexe sur ce plan. Étant donné la difficulté et le coût de ce genre de recherches, il pourrait sembler raisonnable de s’intéresser exclusivement à l’espèce humaine, voire à quelques espèces proches parentes. Cependant, l’approche comparative va faire les preuves de son efficacité. On pourrait, comme Nelson & Platnick (1981), caractériser la biologie générale comme la recherche des invariants à travers le monde vivant, et la biologie comparative comme l’explication des différences au sein de la biodiversité. Il ne semble cependant pas nécessaire d’opposer philosophiquement l’une à l’autre : la diversité est faite de propriétés « générales » à extension plus ou moins limitée. Plus précisément, cet article vise à montrer comment l’étude neurophysiologique d’organismes apparemment « simples » a pu éclairer le fonctionnement d’organismes plus complexes ; et ceci répond bien aux objectifs de la biologie générale. Mais nous verrons également comment l’approche comparative, qui est par excellence l’approche phylo-génétique, peut aider à orienter efficacement le choix de modèles-clés pour la compréhension de l’émergence des capacités cognitives et comportementales au cours de l’évolution.
Évolution et émergence des comportements complexes et des capacités cognitives
2 La vision philosophique du monde (ontologie) implicitement mise en œuvre par les chercheurs en sciences de la nature est matérialiste, évolutionniste, systémiste et émergentiste. Tout ce qui existe pour la science est constitué de systèmes matériels, eux-mêmes composés de sous-systèmes, et en interaction avec d’autres systèmes. Les propriétés émergentes d’un système, que ne possèdent pas ses parties, ne « tombent pas du ciel », elles ne sont pas non plus « produites par » l’interaction des sous-systèmes de façon inexplicable ; ces propriétés sont l’interaction même entre les sous-systèmes, le système étant lui-même en interaction avec son environnement, composé d’autres systèmes matériels (Bunge, 1981, 2008 ; Mahner & Bunge, 1997). Dans la perspective évolutive « d’héritage avec modification » des caractéristiques biologiques, on doit s’attendre à trouver des organismes complexes composés de sous-systèmes plus simples hérités de leur passé évolutif. L’étude de la structure et des lois de fonctionnement d’organismes simples pourrait donc montrer des « systèmes élémentaires » encore présents dans des organismes plus évolués (notion de « conservation des gains »), et qui sont parties prenantes de leurs propriétés émergentes. En d’autres termes, l’étude des systèmes les plus simples ouvre la voie à la compréhension du fonctionnement des systèmes complexes qui les intègrent sous une forme identique ou peu modifiée. L’étude d’espèces animales n’est donc pas uniquement un succédané imposé par les limites éthiques et technologiques de l’expérimentation sur l’espèce humaine, elle constitue une approche réellement efficace pour comprendre cette dernière.
Approche comparative dans l’étude du système nerveux
3 Lorsque l’on s’intéresse au système nerveux en tant que lieu biologique des processus cognitifs complexes présents chez l’homme, il est nécessaire d’étudier tous les niveaux d’organisation depuis les molécules impliquées dans la communication cellulaire et les réseaux neuronaux jusqu’aux comportements, et même les interactions sociales entre les individus. Ces différents niveaux de fonctionnement existent déjà chez des espèces plus simples.
4 On peut étudier le fonctionnement des synapses chez l’homme ou même les vertébrés en général, qui possèdent un système nerveux complexe ; mais cette complexité en rend l’étude difficile. Or, neurones, synapses et réseaux neuronaux existent depuis l’apparition des premiers animaux triploblastiques, comme les cnidaires : hydres et méduses. Par exemple, les gènes impliqués dans le développement des neurones sont déjà présents et peuvent être étudiés chez ces espèces à organisation simple.
5 Le système nerveux ne se constitue que lorsque le troisième feuillet (mésoderme) apparaît au cours de l’évolution. Les neurones sont en effet formés à partir de cellules ectodermiques par interaction avec des cellules mésodermiques. Cependant, l’ectoderme existant déjà chez les animaux diploblastiques, certaines propriétés du système nerveux peuvent y être étudiées. Par exemple, la plupart des gènes de régulation des synapses sont présents chez les éponges, montrant que certaines caractéristiques nerveuses peuvent même être étudiées chez un organisme simple dépourvu d’un authentique système nerveux (Sakarya et al., 2007 ; Richards et al., 2008).
6 Ces exemples montrent qu’il doit être possible de trouver dans des organismes simples des précurseurs de systèmes biologiques complexes.
Réseaux neuronaux
7 Un réseau neuronal est composé d’une multitude de neurones interconnectés et interagissant. La boucle réflexe peut être considérée chez les vertébrés comme l’un des plus simples systèmes neuronaux. Par exemple le réflexe myotatique simple des vertébrés nécessite juste un récepteur sensoriel, un neurone sensoriel, un interneurone inhibiteur et deux neurones moteurs (figure 1). Chez les cnidaires, premiers animaux présentant un système nerveux (Martin & Thomas,1981 ; Westfall et al., 1971), ces réseaux simples, ainsi que les différents types de récepteurs sensoriels impliqués (chémorécepteurs, statorécepteurs et photorécepteurs) sont déjà présents. De plus, ces animaux possèdent aussi des neurones associatifs dans des ganglions et dans un réseau nerveux diffus qui leur permettent un comportement cohérent en relation avec les informations sensorielles. Par exemple, une méduse va pouvoir se déplacer en maintenant son équilibre grâce aux informations en provenance des statorécepteurs qui détectent les variations de position de l’animal par rapport à l’horizontale. Ces informations sont intégrées par un réseau neuronal associatif qui envoie les commandes aux muscles (figure 1). Ces espèces nous offrent ainsi des modèles animaux simples permettant l’étude d’interactions neuronales plus complexes (Koizumi, 2002 ; Shimizu, 2002).
Réflexe myotatique simple des vertébrés (à gauche) où l’étirement d’un muscle extenseur produit l’activation d’un récepteur (fuseau neuromusculaire) et d’un petit réseau neuronal de la moelle épinière qui a pour conséquence la contraction du même muscle extenseur et l’inhibition du muscle antagoniste. Chez la méduse (à droite) le balancement des statocystes (récepteurs de position) active un réseau neuronal qui permet à l’animal de garder son équilibre et ainsi d’avoir un déplacement cohérent.

Réflexe myotatique simple des vertébrés (à gauche) où l’étirement d’un muscle extenseur produit l’activation d’un récepteur (fuseau neuromusculaire) et d’un petit réseau neuronal de la moelle épinière qui a pour conséquence la contraction du même muscle extenseur et l’inhibition du muscle antagoniste. Chez la méduse (à droite) le balancement des statocystes (récepteurs de position) active un réseau neuronal qui permet à l’animal de garder son équilibre et ainsi d’avoir un déplacement cohérent.
Les processus de développement du système nerveux et de mise en place des réseaux neuronaux peuvent être étudiés chez des organismes simples
8 Les organismes simples possédant ces différents types de neurones présentent également un développement rapide et des capacités régénératives importantes. Ils peuvent donc nous permettre aussi d’étudier des processus développementaux et plus largement la plasticité neuronale : apparition des cellules nerveuses, formation des contacts synaptiques et des molécules nécessaires à la transmission de l’influx nerveux. Les capacités régénératives de ces organismes primitifs sont telles qu’ils peuvent nous permettre aussi d’étudier ces processus développementaux lors de régénérescences de parties lésées (Koizumi, 2002). Si la tête de l’hydre est sectionnée, elle se régénère totalement, ainsi que toutes ses structures internes et ses réseaux neuronaux complexes. Cette régénérescence, complètement réalisée en moins de 4 jours, fait de cet animal un excellent modèle d’étude du développement du système nerveux et de la plasticité neurale en général.
Système nerveux, comportement et cognition
9 Les exemples précédents nous ont montré l’existence de tous les éléments de bases nécessaires à l’existence d’un système nerveux dans des organismes relativement simples. Ces systèmes nerveux simples sont à la base des comportements simples de ces espèces. Si l’on s’intéresse aux capacités cognitives complexes de l’homme, on peut supposer l’existence des bases cognitives chez des espèces plus simples.
10 Pour tous types d’apprentissages, les capacités de mémorisation à court et long terme sont indispensables ; et ces capacités existent déjà chez la plupart des invertébrés. L’aplysie, un mollusque gastéropode marin, a été un modèle clé pour comprendre les processus neuronaux impliqués dans la mémorisation à court et long terme (Kandel & Schwartz, 1982 ; Kandel et al., 1987).
11 Chez l’aplysie le comportement simple de retrait du siphon et des branchies lors d’une stimulation tactile montre les capacités d’apprentissage chez cet animal (Kandel, 1976). Quand on touche le siphon à l’aide d’un pinceau, il y a retrait du siphon et des branchies. Une dizaine de stimulations provoquent une habituation à court terme d’une dizaine de minutes. Si on répète cet exercice sur plusieurs jours (4 en l’occurrence), cette habituation dure trois semaines et l’on parle d’habituation à long terme.
12 Le mécanisme est connu : la stimulation des interneurones facilitateurs du circuit de rétraction des branchies leur fait libérer de la sérotonine sur les terminaisons présynaptiques des neurones sensoriels innervant le siphon. Ces derniers libèrent alors davantage de neurotransmetteur à leur synapse avec les neurones moteurs de la branchie, provoquant ainsi une sensibilisation du système (Kandel et al., 1987).
La fonction de l’hippocampe chez les vertébrés supérieurs
13 Les travaux sur l’aplysie ont facilité l’étude de l’hippocampe, structure cérébrale impliquée dans les processus de mémorisation chez les vertébrés supérieurs. Des expériences de lésion de l’hippocampe chez l’animal, ainsi que des données cliniques chez l’homme, ont montré son importance dans les processus de mémorisation et d’apprentissage en général. Par exemple, dans un apprentissage spatial l’intégrité de l’hippocampe est indispensable. Un rat placé dans une piscine remplie d’eau trouble est capable de trouver une plate-forme cachée sous le niveau de l’eau. Après une dizaine d’essais, il aura appris à nager directement vers cette plate-forme. Un rat présentant des lésions dans l’hippocampe ne sera pas capable d’apprendre cette tâche (Schenk & Morris, 1985). Chez l’homme, des lésions de l’hippocampe induisent des déficits de mémorisation tels que les patients ne sont plus capables de nouveaux apprentissages alors qu’ils se souviennent d’événements anciens (Scoville & Milner, 1957 ; Hilts, 1995). L’étude de l’hippocampe a montré l’existence de circuits neuronaux comparables à ceux décrits chez l’aplysie, dans lesquels des stimulations répétées ou simultanées induisent des potentialisations à long terme (Nicoll et al., 1988). Bien que les neurotransmetteurs utilisés par ces circuits soient différents, l’organisation neuronale, qui est du même ordre, montre là aussi que le substrat biologique de la mémorisation chez l’homme peut être étudié chez des modèles animaux plus simples puisque ce substrat existe déjà chez ces espèces.
Langage chez l’homme et chant chez les oiseaux chanteurs
14 Nous venons de voir qu’il est relativement aisé de trouver des modèles animaux simples pour étudier la physiologie du système nerveux mais aussi les bases neurales de comportements et de capacités cognitives relativement complexes. Cependant, lorsqu’on tente de trouver un modèle animal pour étudier le langage humain cela devient plus difficile. Il est vrai que l’on s’attaque là à ce qui est supposé être « le propre de l’homme », c’est-à-dire l’une des capacités cognitives qui le différencie de tous les autres animaux et qui donc, par définition, ne peut pas être retrouvée chez d’autres espèces. Tout se passe comme si le langage était une capacité cognitive qui n’apparaît que chez l’homme. Si l’on cherche cette capacité chez des espèces phylo-génétiquement proches on s’aperçoit qu’aucun primate non humain ne possède de langage comparable à celui des primates humains. Cela est vrai si l’on ne cherche cette capacité que parmi les mammifères. Or, si l’on cherche ces capacités dans d’autres classes animales, on va s’apercevoir que chez certaines espèces d’oiseaux qui utilisent leurs vocalisations pour communiquer, les chants sont variés et présentent une syntaxe complexe. La complexité n’atteint bien évidemment pas celle du langage humain et les informations véhiculées par ces signaux sont beaucoup plus simples, mais nous avons là des espèces animales qui peuvent nous permettre d’étudier les facteurs pouvant influencer l’apparition du langage. C’est ainsi que depuis les années 1960 le chant des oscines (« oiseaux chanteurs » parmi les passereaux) est devenu un bon modèle animal d’apprentissage du langage.
15 Il y a quelques similitudes entre langage humain et chants d’oiseaux. Les oscines apprennent à chanter en passant par différents stades comparables à ceux connus pendant l’apprentissage du langage chez l’homme (Marler, 1970). De plus, comme l’homme et de nombreux mammifères, ces oiseaux utilisent leurs vocalisations dans leurs interactions sociales. Ils ont été longtemps considérés comme un exemple de convergence, des espèces éloignées développant des comportements analogues pour remplir les mêmes fonctions. Les progrès en génétique du développement ont montré qu’il ne s’agissait pas d’une analogie mais d’une véritable homologie des structures impliquées (figure 2). Les mêmes structures cérébrales sont utilisées chez l’homme et les oscines pour percevoir, traiter et produire des signaux vocaux de communication (Jarvis et al., 2004).
16 Parmi ces oiseaux chanteurs, l’étourneau sansonnet possède des capacités cognitives qui en font un modèle pour l’étude des apprentissages vocaux. Comme tous les oscines il apprend son chant spécifique en présence d’un tuteur adulte mais surtout en l’écoutant et en interagissant avec lui. Cependant, contrairement à la plupart des oscines, il ne subit pas une cristallisation de son chant en fin d’apprentissage, ce qui lui donne la possibilité d’apprendre de nouvelles vocalisations tout au long de sa vie. Cette capacité lui permet d’enrichir son répertoire vocal et de donner une dimension individuelle à ses vocalisations (Hausberger et al., 2000). Cela en fait un excellent modèle animal pour étudier le langage, son apprentissage et son utilisation.
Cerveau d’oiseau (à gauche) et e mammifère (à droite). A - Jusqu’au début des années 2000 il était admis que la majeure partie du télencéphale des oiseaux était constituée de striatum et il était donc considéré comme plus primitif que le cerveau des mammifères et très différent de ce dernier. B - Les récents progrès en génétique du développement ont montré que les structures nerveuses constituant le télencéphale des oiseaux ont la même origine palliale que chez les mammifères. Les mêmes structures palliales sont impliquées dans des tâches cognitives complexes comparables telles que les traitements des signaux de communication
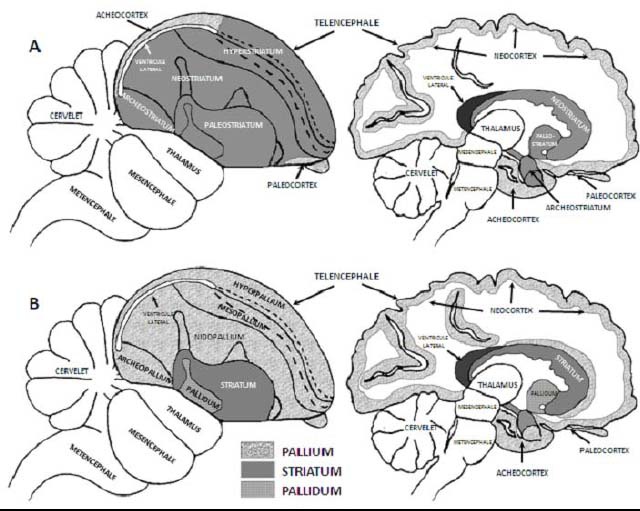
Cerveau d’oiseau (à gauche) et e mammifère (à droite). A - Jusqu’au début des années 2000 il était admis que la majeure partie du télencéphale des oiseaux était constituée de striatum et il était donc considéré comme plus primitif que le cerveau des mammifères et très différent de ce dernier. B - Les récents progrès en génétique du développement ont montré que les structures nerveuses constituant le télencéphale des oiseaux ont la même origine palliale que chez les mammifères. Les mêmes structures palliales sont impliquées dans des tâches cognitives complexes comparables telles que les traitements des signaux de communication
17 Des expérimentations menées sur cet animal ont montré d’une part, l’importance de l’expérience individuelle précoce en présence de congénères adultes et d’autre part, la nécessité d’interactions sociales avec ces adultes pour développer non seulement la production du chant mais aussi la capacité à percevoir ce chant (Cousillas et al., 2004, 2006).
18 Des animaux isolés socialement ne développent pas l’organisation fonctionnelle cérébrale adéquate pour pouvoir traiter les signaux de communication de leur espèce (Cousillas et al., 2008). Ces résultats, obtenus sur un modèle simple, et décrivant l’importance des interactions sociales dans un apprentissage vocal, présentent des analogies claires avec l’apprentissage du langage chez l’homme. Certains facteurs influençant l’apprentissage vocal chez l’homme pourraient donc être étudiés plus facilement sur ce modèle animal.
Un aperçu des méthodes
19 Les méthodes utilisées pour faire ces études sont essentiellement basées sur des enregistrements électrophysiologiques faits en général à l’aide de microélectrodes. Ce type d’enregistrements donne une précision spatio-temporelle parfaite. Sur le plan spatial nous sommes au niveau de la membrane cellulaire, de la cellule ou au plus de quelques cellules selon le type de microélectrodes. Sur le plan temporel la précision peut être de l’ordre de la microseconde, permettant d’affirmer à quel moment s’est produit le phénomène observé. Cela nous a permis par exemple, à l’aide d’enregistrements multicellulaires et unitaires effectués à intervalles réguliers (figure 3), de faire une cartographie des aires activées pendant la diffusion de signaux de communication (Hausberger et al., 2000 ; Cousillas et al., 2005).
Méthode d’enregistrement électrophysiologique permettant la production de cartes d’activité. Représentation schématique d’un cerveau d’oiseau chanteur (à gauche) avec les structures impliquées dans la perception auditive (en rouge), la production vocale (en bleu) et l’apprentissage vocal (en vert). L’exemple présenté ici concerne des enregistrements effectués à intervalles réguliers (au milieu) pendant la diffusion de vocalisations dans le Champ L du cerveau d’étourneau (homologue du cortex auditif humain). Après compilation de tous les résultats une carte d’activité est construite (à droite). Le code de couleur permet de montrer le taux d’activation des neurones par rapport à l’activité spontanée (activité de base = 100 %).
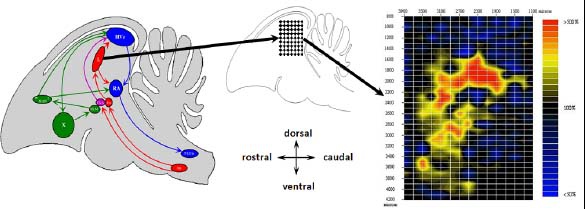
Méthode d’enregistrement électrophysiologique permettant la production de cartes d’activité. Représentation schématique d’un cerveau d’oiseau chanteur (à gauche) avec les structures impliquées dans la perception auditive (en rouge), la production vocale (en bleu) et l’apprentissage vocal (en vert). L’exemple présenté ici concerne des enregistrements effectués à intervalles réguliers (au milieu) pendant la diffusion de vocalisations dans le Champ L du cerveau d’étourneau (homologue du cortex auditif humain). Après compilation de tous les résultats une carte d’activité est construite (à droite). Le code de couleur permet de montrer le taux d’activation des neurones par rapport à l’activité spontanée (activité de base = 100 %).
20 Ces méthodes sont facilement utilisables sur des modèles animaux simples et très difficilement, voire même inutilisables, chez l’homme. Ces dernières années, les grands progrès en imagerie cérébrale ont révolutionné les études fonctionnelles du cerveau chez l’homme. Ces méthodes ont permis de mettre en évidence des zones cérébrales qui s’activent lors de tâches particulières. Lors de tests impliquant des tâches langagières, les aires cérébrales qui avaient été caractérisées auparavant à l’aide de données cliniques ont été activées. Par exemple l’aire de Broca connue pour son implication dans la production du langage s’active lorsque le sujet pense à un verbe. Ces techniques ont permis de grandes avancées en clinique bien sûr mais aussi en recherche humaine. Cependant, la précision spatio-temporelle (au mieux, la taille des aires cérébrales sur le plan spatial, et de l’ordre de quelques secondes sur le plan temporel) n’est pas suffisante pour étudier l’organisation fonctionnelle d’un réseau neuronal. Bien que comparativement trop imprécise, l’identification des aires cérébrales activées peut cependant être d’une grande utilité car ces techniques donnent une image d’ensemble qui nous permet de localiser toutes les zones cérébrales activées lors d’une tâche particulière. Ces mêmes techniques peuvent être utilisées chez l’animal pour obtenir une image d’ensemble des zones activées lors de tâches cognitives. Chez les oiseaux chanteurs les zones impliquées dans le traitement et la production des signaux vocaux de communication peuvent être mises en évidence par ces techniques d’imagerie (figure 4). Ces résultats nous permettraient ensuite de cibler de manière précise les aires cérébrales où nous pourrons enregistrer les neurones par microélectrodes (méthode trop invasive pour être utilisée chez l’homme), ouvrant ainsi la voie à l’étude de l’organisation fonctionnelle d’un petit réseau neuronal impliqué dans une tâche cognitive complexe.
Cerveau d’étourneau observé en IRM anatomique (coupe sagittale à gauche, horizontale à droite). Cette technique permet d’identifier et de caractériser les différentes structures du cerveau. Cela permet par exemple de vérifier la position des électrodes d’enregistrement, ce qui évite le prélèvement du cerveau pour observation en histologie. L’association de l’IRM fonctionnelle à ces images, comme cela est fait dans l’équipe d’A. Van der Linden à Anvers (Van Meir et al., 2005), permet de repérer les zones activées par des vocalisations spécifiques, et de mieux cibler ensuite les zones à enregistrer par microélectrodes.
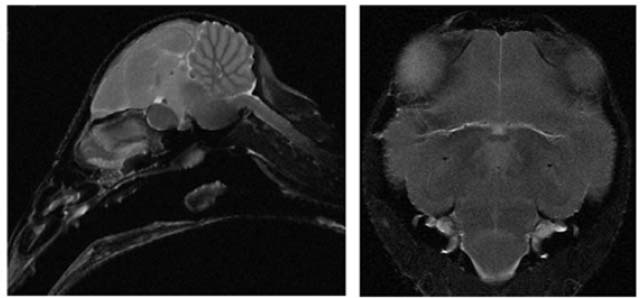
Cerveau d’étourneau observé en IRM anatomique (coupe sagittale à gauche, horizontale à droite). Cette technique permet d’identifier et de caractériser les différentes structures du cerveau. Cela permet par exemple de vérifier la position des électrodes d’enregistrement, ce qui évite le prélèvement du cerveau pour observation en histologie. L’association de l’IRM fonctionnelle à ces images, comme cela est fait dans l’équipe d’A. Van der Linden à Anvers (Van Meir et al., 2005), permet de repérer les zones activées par des vocalisations spécifiques, et de mieux cibler ensuite les zones à enregistrer par microélectrodes.
Conclusion
21 Nous avons montré chez différentes espèces animales l’existence d’une unité structurale et fonctionnelle dans le système nerveux. Tous les éléments de bases constituant le système nerveux existent déjà chez des organismes simples. Les cellules de base (neurones et cellules gliales), ainsi que les réseaux neuronaux, sont présents chez tous les animaux possédant un système nerveux et les gènes impliqués dans le fonctionnement des neurones peuvent même être étudiés chez des animaux dépourvus de système nerveux. Cela se passe un peu comme si la complexification du système nerveux au cours de l’évolution consistait simplement en une combinaison de sous-systèmes plus simples. Un cerveau complexe serait ainsi composé de nombreux sous systèmes qui peuvent être retrouvés chez des organismes plus simples et donc être étudiés chez ces derniers. Les capacités cognitives et les comportements complexes en général sont une émanation du système nerveux et permettent à l’individu de percevoir et d’agir sur son environnement et sur ses congénères. Si, comme pour les sous-systèmes composants le système nerveux, les bases comportementales fonctionnelles sont présentes chez des animaux simples, l’étude phylogénique des comportements peut être d’une grande utilité pour comprendre le comportement humain. La communication vocale atteint avec le langage humain à une complexité inégalée dans le règne animal. Nous avons vu qu’en raison des similitudes entre langage et chant d’oiseau, il était possible d’étudier quelques règles qui régissent l’apprentissage du langage sur le modèle plus simple des oiseaux chanteurs. Il serait intéressant d’aller plus loin et d’essayer de connaître les origines de la capacité à vocaliser et surtout à utiliser ces vocalisations pour communiquer et interagir socialement. La phylogénie est essentielle pour choisir les taxons à étudier en priorité. Parmi les amniotes, la communication vocale est bien développée dans deux classes, les oiseaux et les mammifères, avec une sophistication particulière chez les oiseaux chanteurs et les primates humains. Chez les reptiles, la plupart des espèces ne vocalisent pas mais les crocodiliens interagissent vocalement (Vergne et al., 2009). Or, oiseaux et crocodiles appartiennent au même phylum, les archosauriens. L’étude chez les crocodiliens des vocalisations ainsi que des bases neurales de leur perception et de leur production pourrait nous fournir des connaissances essentielles sur les fondements de la capacité à vocaliser et à communiquer par cette modalité. La comparaison avec les autres reptiles « muets » nous informerait sur l’apparition de cette capacité cognitive dans ce phylum. Ces études peuvent aussi apporter un éclairage nouveau sur l’évolution menant aux oiseaux, déjà démontrés similaires aux mammifères sur plusieurs plans.
22 Plus généralement, selon le comportement et les capacités cognitives auxquels nous nous intéressons, nous pouvons nous appuyer sur la phylogénie pour déterminer les organismes cibles dont on peut privilégier l’étude pour développer l’approche comparative la plus prometteuse.
Références
- Bunge M. 1981. Scientific materialism. D. Reidel, Dordrecht.
- Bunge M. 2008. Le matérialisme scientifique. Traduit de l’anglais par S. Ayache, P. Deleporte, E. Guinet et J. Rodriguez-Carvajal. Syllepse, Paris.
- Hausberger M, Leppelsack E, Richard J.-P. & Leppelsack H.J. 2000. Neuronal bases of categorization in starling song. Behav. Brain Res. 114, 89-95.
- Hilts, P.J. 1995. Memory’s ghost : the strange tale of Mr. M and the nature of memory. Simon and Schuster, New York.
- Mahner M. & Bunge M. 1997. Foundations of biophilosophy. Springer-Verlag, Berlin.
- Nelson G. & Platnick N. 1981. Systematics and biogeography : cladistics and vicariance. Columbia University Press, New York.
- Jarvis E. D., Gþntþrkþn O. & Bruce L. 2005. Avian brains and a new understanding of vertebrate brain evolution. Nature Reviews - Neuroscience 6, 151-159.
- Jarvis E.D. 2004. Learned birdsong and the neurobiology of human language. Ann. NY Acad. Sci. 1016, 749-777.
- Kandel E.R. 1976. Cellular basis of behavior. W.H. Freeman, San Francisco.
- Kandel E.R. & Schwartz J.H. 1982. Molecular biology of learning : modulation of transmitter release. Science 218, 433-443.
- Kandel E.R., Castelluci V.F., Goelet P. & Schacher S. 1987. Cell-biological interrelationships between short-term and long-term memory. Ass. for Res. in Nervous and Mental disease 65, 11-132.
- Koizumi O. 2002. Developmental neurobiology of Hydra, a model animal of cnidarians. Can. J. Zool., 80, 1678-1689.
- Marler P. 1970. Birdsong and speech development : could there be parallels ? Am. Sci. 58, 669-673.
- Martin V.J. & Thomas M.B. 1981. The origin of the nervous system in Pennaria Tiarella, as revealed by treatment with colchicines. Biol. Bull. 160, 303-310.
- Nicoll R.A., Kauer J.A. & Malenka R.C. 1988. The current excitement in long term potentiation. Neuron 1, 97-103.
- Richards G.S., Simionato E., Perron M., Adamska M., Vervoort M. & Degnan B. 2008. Sponge genes provide new insight into the evolutionary origin of the neurogenic circuit. Current Biol. 18, 1156-1161.
- Sakarya O., Armstrong K.A., Adamska M., Adamski M., Wang I-F., Tidor B, Degnan B.M., Oakley T.H. & Kosik K.S. 2007. A post-synaptic scaffold at the origin of the animal kingdom. PLos ONE 6, e506.
- Schenk F. & Morris R.G.M. 1985. Dissociation between components of spatial memory in rats after recovery from the effects of retrohippocampal lesions. Exp. Brain Res. 58, 11-28.
- Scoville W.B. & Milner B. 1957. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 20, 11-21.
- Shimizu H. 2002. Feeding and wounding responses in Hydra suggest functional and structural polarization of the tentacle nervous system. Comp. Biochem. and Physiol. 131, 669-674.
- Vergne A.L., Pritz M.B. & Mathevon N. 2009. Acoustic communication in crocodilians : from behaviour to brain. Biol. Rev. 84, 391-411.
- Westfall J.A., Yamataka S. & Enos P.D. 1971. Ultrastructural evidence of polarized synapses in the nerve net of hydra. J. Cell Biol. 51, 318-323.