Classification et tempérament chez le chien domestique (Canis familiaris) : sélection naturelle et artificielle de caractères morphologiques et comportementaux
Pages 145 à 160
Citer cet article
- DEPUTTE, Bertrand L.
- et DELEPORTE, Pierre,
- Deputte, Bertrand L..
- et al.
- Deputte, B.-L.
- et Deleporte, P.
https://doi.org/10.3917/biosy.027.0145
Citer cet article
- Deputte, B.-L.
- et Deleporte, P.
- Deputte, Bertrand L..
- et al.
- DEPUTTE, Bertrand L.
- et DELEPORTE, Pierre,
https://doi.org/10.3917/biosy.027.0145
Introduction
1 La systématique et la taxinomie peuvent se concevoir à tous les degrés de l’organisation et de l’histoire du vivant, mais les approches au niveau des populations ou des races domestiques demeurent des parents pauvres de la réflexion dans ce domaine, a fortiori en ce qui concerne les caractères éthologiques. L’objet de cet article est de présenter les hypothèses actuelles quand à l’origine des caractéristiques comportementales des chiens domestiques, d’examiner la pertinence des standards raciaux concernant les traits de tempérament des chiens actuels, et d’évoquer des voies de recherche possibles pour une amélioration de leur sélection artificielle prenant en compte le tempérament.
Comportement et tempérament : définitions
2 Le comportement peut se définir comme une manifestation motrice exprimée par un animal à la suite de la perception d’informations du milieu extérieur et de l’état interne de son corps (p. ex. Manning, 1972 ; Campan & Scapini, 2002). Le tempérament fait référence aux différences interindividuelles comportementales entre individus placés dans des situations similaires. Le tempérament représente une caractéristique individuelle, multidimensionnelle, stable dans le temps et entre les situations. Il est génétiquement influencé, apparaît précocement et fait référence à un substrat biologique (Goldsmith, 1983 ; Goldsmith et al., 1987). Le tempérament représente en partie ce que l’on peut appeler la « structure émotionnelle de l’individu », c’est-à-dire ce qui déterminera un individu à réagir de manière prédictible dans des situations potentiellement anxiogènes (apparition de stimulus soudain, placement dans un environnement nouveau, mise en présence d’objets nouveaux, isolement social, capacité à apprendre, etc. ; Svartberg & Forkman, 2002 ; Jones & Gosling, 2005). Le tempérament inclut également le niveau d’activité de l’individu, son degré de motivation et de persévérance dans des tâches spécifiques. Le tempérament est constitué de nombreux traits correspondant à ses différentes dimensions. L’un des gradients le plus souvent mentionnés est celui de « Timidité/Intrépidité » (Shyness-Boldness ; Kagan et al., 1988 ; Wilson et al., 1993 ; Svartberg & Forkman, 2002). Il fait écho à la théorie biphasique du développement « Approche-Evitement » (Approach-Withdrawal) proposée par Schneirla (1959) qui stipule, notamment, qu’un organisme dans les stades précoces de son développement aura tendance à approcher des stimuli de faible intensité et à s’éloigner de stimuli de forte intensité.
3 Le comportement est directement observable, le tempérament est une inférence faite à partir des comportements observés dans différentes situations particulières.
Évolution et sélection du comportement des chiens domestiques
4 La sélection naturelle conduit à la différenciation d’espèces dont on ne peut que constater a posteriori l’adaptation à un certain milieu. Des atteintes, notamment anthropiques, à ces milieux posent des défis permanents à ces espèces. On ne peut, pour ces espèces sauvages, que spéculer sur les facteurs de sélection qui ont conduit à façonner les espèces présentes. Mais d’autres espèces, bien moins nombreuses, ont connu ou connaissent encore, en plus de leur sélection naturelle, une sélection artificielle conduite par l’homme dans une ou plusieurs directions explicites. C’est le cas des espèces domestiquées, dont la grande diversité, les variations et leur conservation, ont inspiré à Darwin sa théorie de l’évolution. Parmi les espèces domestiques, le chien tient une place particulière. La spéciation du chien à partir de l’espèce-mère, le loup gris, a donné lieu à l’élaboration de plusieurs scénarios. Clutton-Brock (1999), suivant l’hypothèse de Darwin, met l’accent sur un rôle des humains très précoce au cours du processus de sélection artificielle, en affirmant « we cannot know when human hunters first tamed wolves » (Clutton-Brock, 1999 : 56). Toutefois elle souligne que la domestication du chien relève d’un double processus, le premier, très lent, de sélection naturelle de l’espèce-mère, le deuxième, beaucoup plus rapide, d’une sélection artificielle appliquée par l’homme (Clutton-Brock, 1995). Envisager un apprivoisement précoce de loups « sauvages » impliquerait qu’au Paléolithique supérieur de nombreux groupes d’Homo sapiens encore nomades auraient attiré à eux un prédateur ou se seraient emparés de jeunes pour les faire élever en leur compagnie. Le résultat en aurait été une modification du comportement de ces carnivores sauvages rappelant l’expérience de Belayev sur les renards argentés (Trut, 1999). Ce généticien russe a pendant quarante ans sélectionné systématiquement parmi des renards captifs élevés pour leur fourrure ceux qui présentaient la plus grande attraction spontanée vers l’humain. Cependant cette sélection artificielle, explicite et systématique, a produit un nombre significatif mais relativement restreint de sujets possédant ce trait par contraste avec le reste de l’espèce, et en outre les individus ainsi produits n’avaient plus aucun contact avec la population sauvage ou avec les renards argentés captifs qui ne possédaient pas ce trait.
5 La domestication du chien aurait-elle pu suivre un processus similaire ? Dans une étude par modélisation sur l’ADN nucléaire (diversité allélique des gènes du complexe majeur d’histocompatibilité), Vilà et al. (1997) proposent deux dates pour la divergence génétique entre le loup et le chien : 135 000 BP et 15 000 BP. La première date poserait la question de savoir si c’est Homo sapiens ou Homo neanderthalensis qui aurait débuté la « domestication » du chien. La deuxième, qui selon Vilà et al. (1997) correspondrait à l’apparition d’une différenciation morphologique entre le loup-loup et le loup-chien, se situe clairement dans l’époque d’Homo sapiens, Homo neanderthalensis ayant alors disparu. Une question demeure : l’Homo sapiens du Paléolithique supérieur avait-il déjà les connaissances de Belyaev en matière de sélection ? Peut-on envisager une divergence si profonde par la répétition d’événements isolés d’apprivoisement avec des individus qui ne seraient pas maintenus séparés des individus sauvages apprivoisés ? Coppinger et Coppinger (2001 : 41) proposent un scénario qui se résume dans cette phrase : « I think a population (at least one) of wolves domesticated themselves. » Ce scénario met l’accent sur le processus de sélection naturelle qui serait essentiellement responsable de la spéciation du chien à partir du loup. Ce scénario repose en partie sur le constat fait par ces auteurs que de par le monde, il existe une majorité de chiens, morphologiquement bien différents des loups, qui vivent en situation de commensaux de l’homme, c’est-à-dire complètement libres de réguler leurs proximités et leurs distances avec les humains sans être ni captifs ni semi-captifs. Ce scénario s’accorde avec l’ensemble des dates fournies par les différents auteurs ayant travaillé sur la phylogenèse moléculaire des chiens et des loups (Vilà et al., 1997 ; Savolainen et al., 2002). Il s’accorde aussi avec le fait que cette spéciation, ou du moins cette divergence phénotypique, se soit produite à une grande échelle et dans des zones géographiques relativement larges. Il envisage donc que des groupes de loups se soient approchés des campements humains probablement pour tirer profit des restes alimentaires. Cette hypothèse postule que des loups habituellement compétiteurs de l’humain, se disputant les mêmes ressources alimentaires, se soient montrés plus intrépides que d’autres pour approcher de ces chasseurs bipèdes efficaces. Ce trait de tempérament aurait donc pu être l’élément déterminant dans une radiation au sein de l’espèce Canis lupus. Posséder une double compétence comportementale, soit de pouvoir chasser des proies seul ou en groupe, soit de pouvoir profiter de ressources acquises par une espèce sympatrique et prédatrice efficace, offrirait de toute évidence un avantage sélectif, qui aurait pu conduire à une divergence de plus en plus marquée entre les loups-timides et les loups-intrépides. Ce trait de tempérament a une grande probabilité d’apparaître dans de nombreux groupes de loups sympatriques d’humains, donc de créer la possibilité d’une radiation sur une large échelle. Cette propagation peut paraître plus probable que celle de la décision d’un apprivoisement de loups par des humains. Cette hypothèse a aussi l’attrait de proposer un scénario plausible alors même que l’apprivoisement de loups par les humains avec pour seule finalité d’apprivoiser des carnivores compétiteurs paraît évolutivement moins plausible. Le scénario d’une spéciation du chien par sélection naturelle d’un trait de tempérament s’accorde avec les résultats d’études génétiques postulant que les chiens sont originaires soit de plusieurs populations et de centaines d’individus, soit d’un plus petit nombre de fondateurs qui aurait progressivement augmenté par croisement avec des loups (Vilà et al., 2005). Cette dernière hypothèse serait cependant moins en accord avec le trait de tempérament envisagé.
6 La grande diversité phénotypique actuelle, avec 330 races reconnues par la Fédération cynologique internationale (FCI) (Anonyme, 2009), est relativement récente et date de l’époque victorienne, celle de Darwin. Cette diversité affecte les traits morphologiques et les traits comportementaux, deux catégories souvent non indépendantes et en liaison avec une sélection explicite des humains mieux dotés culturellement que ceux du Paléolithique supérieur. Le chien pourrait donc provenir de la succession de deux phases de sélection de caractères particuliers. La première phase relève d’une sélection naturelle avec la sélection d’un trait de tempérament, et la deuxième phase probablement de plusieurs épisodes de sélection artificielle dont l’apogée a été observé au XIXe siècle. C’est cette deuxième phase qui mérite le terme de domestication du chien, car comme le souligne Franck (1980), « any domestic species is almost by definition a product of artificial selection ».
7 La sélection artificielle du chien a de toute évidence modelé son comportement en fonction des tâches qui lui étaient assignées. Coppinger & Coppinger (2001) ont proposé un scénario possible de cette sélection artificielle pour les chiens de chasse et de garde ou de conduite de troupeau. Ils envisagent, à partir d’une séquence de patterns moteurs de prédation chez un canidé sauvage, différentes modalités d’hypertrophie ou d’inhibition de ces patterns moteurs chez différentes races domestiques (Coppinger & Coppinger, 2001 : 199). Selon leur scénario, quasiment l’ensemble de la séquence comportementale aurait été préservé au cours de la sélection de chiens courants alors qu’à l’autre extrême toute la séquence aurait été inhibée chez les chiens de garde de troupeau, comme les grandes races de montagne. Parmi les intermédiaires les plus spectaculaires, ils citent d’une part, les chiens de conduite de troupeau, chez lesquels toute la séquence initiale de localisation, d’approche et de poursuite de la proie serait conservée de manière « hypertrophiée », et le reste inhibé, et d’autre part les « pointers » chez lesquels seuls les patterns moteurs initiaux de la séquence, « s’orienter » et « fixer », et un des patterns terminaux, « Saisir dans les mâchoires », ont été hypertrophiés. Le cas du chien de compagnie relèverait d’une inhibition totale de la prédation au cours d’une sélection explicite comme cela pourrait être le cas pour le Cavalier King Charles. Toutefois l’engouement pour le chien dans différentes civilisations et cultures a conduit à ce que, notamment en France, tout chien, quelle qu’en soit la race, peut accéder au statut de chien de compagnie. On peut alors facilement considérer que ce qui a été minutieusement sélectionné au cours de centaines de générations ait du mal à s’exprimer dans le contexte d’un appartement ou de sorties en ville, par un animal tenu en laisse.
Classification des races domestiques et standards de tempérament
8 La sélection artificielle exercée sur le chien s’est attachée à produire une combinaison de caractères morphologiques et comportementaux. Scott & Fuller (1965) ont montré que les différences comportementales entre races (5 races pures) étaient bien réelles et importantes et surtout bien fixées génétiquement.
9 Certains caractères comportementaux, comme il en a été fait mention, sont en relation avec la prédation, d’autres avec des traits de tempérament ou des réactions émotionnelles (Scott & Fuller, 1965). La question est de savoir si ces traits de tempérament, qui sont des caractéristiques individuelles ayant un certain degré de déterminisme génétique, peuvent être considérés comme des traits raciaux. En effet des « standards » morphologiques précis, ou certains standards comportementaux (le « pointer » des pointers…) permettent d’assigner un individu à une race bien déterminée. Peut-il en être de même de caractères tels que la propension pour un individu à réagir toujours d’une manière prévisible dans une diversité de situations ? Les standards comportementaux ont été édictés par la FCI, afin de préciser quels traits de tempérament doivent être présents chez tous les individus d’une race donnée (Anonyme, 2009 ; tableau 1). Ces traits sont repris dans de nombreuses publications destinées au grand public pour l’identification de races, essentiellement sur des critères morphologiques (p. ex., Alderton, 2002). En ce qui concerne les Spitz et les chiens nordiques de chasse et de traineau, on peut relever pour 9 races de ce groupe de chiens, 32 traits de tempérament établis par la FCI comme standards comportementaux de ces races, dont certaines ne sont représentées dans le monde que par un petit nombre de sujets (p. ex. Chien d’ours de Carélie, Chien norvégien de macareux, etc. ; Breton, 2008). La lecture de ces « standards » de caractère est très instructive. La FCI reconnaît 330 races et présente pour chacune d’elle une rubrique « Caractère » avec les autres éléments définissant une race. On doit alors penser que ces traits de « caractère » sont aussi caractéristiques de ces races. Dans le tableau 1, on remarque que pour les races Chien courant grec et les Basenjis, chiens primitifs africains, aucun standard de caractère n’est donné. Cela peut paraître surprenant pour le Basenji si l’on se rappelle que cette race faisait partie des 5 races pures que Scott & Fuller (1965) ont étudiées en détail dans leur analyse très complète de la relation entre génétique et comportement social. Un grand nombre de dimensions comportementales a donc été scientifiquement analysé et comparé entre races pures, et la FCI n’a apparemment pas fait référence à ce travail. On note également, au moins pour le Rhodesian Ridgeback, une mention de prudence sur l’uniformité du caractère entre les individus d’une même race, notamment placés dans des cultures différentes. Il faut rappeler que le Rhodesian Ridgeback est un grand chien puissant utilisé dans la chasse des Africains par les colons en Rhodésie (« travail sur civils, utilisations policières » ; tableau 1). On pourrait aussi invoquer un manque de cohérence dans la connaissance du comportement du chien entre la FCI et la législation française sur les races de chiens « catégorisés » comme potentiellement dangereux. En effet les Tosa Inu et les Rotweiler sont placés dans les catégories à risque par la loi alors que le standard de caractère de la FCI ne fait mention d’aucun danger particulier. En revanche, la FCI et la loi s’accordent pour ne pas considérer le labrador comme une race dangereuse et la FCI mentionne que Le Labrador est « Digne de confiance, obéissant, doux envers les enfants. Bon caractère, très agile. […] Intelligent, ardent et docile, il ne demande qu’à faire plaisir. Naturel amical, sans aucune trace d’agressivité ». L’avertissement donné par la FCI pour le Rhodesian Ridgeback aurait cependant dû être appliqué au Labrador car c’est la race la plus impliquée (en données corrigées de l’abondance de la race) dans les morsures domestiques, notamment sur les enfants. Les adverbes utilisés dans les standards raciaux de « caractère » laissent supposer une absence d’analyses scientifiques dans lesquelles les paramètres de dispersion des variables statistiques sont rarement nuls (Basset Hound : « jamais agressif » ; Cavalier King Charles, Labrador : « aucune tendance à… » ; Spitz finlandais : « jamais vicieux »…). Des caractéristiques comme « le désir de plaire » ou « de faire plaisir » sont des notions difficiles à évaluer par des éthologistes (Labrador, Husky, etc.). Pour quelques races, notamment de berger, la FCI souligne à juste titre que la sélection artificielle primaire qui a été à l’origine de ces races de travail est très susceptible de perdurer quelle que soit la nouvelle fonction assignée au chien, notamment celle de compagnie (Berger de Beauce, Border Collie ; tableau 1).
10 On peut se demander si la détermination de ces traits comportementaux considérés comme caractéristiques des races a fait l’objet de tests spécifiquement conçus pour évaluer la présence de ces traits et leur stabilité au fil des générations successives, et quel est le pouvoir discriminant de ces traits considérés comme des standards raciaux comportementaux. Il est difficile d’envisager que les qualificatifs comportementaux ou tempéramentaux de la FCI puissent référer à des phénotypes définis de manière précise. En l’absence de cette précision, il est impossible d’envisager la recherche du support génétique grâce auquel une sélection efficace pourrait être opérée.
11 Récemment, Svartberg & Forkman (2002) ont entrepris une analyse à grande échelle sur les traits de tempérament (qu’ils ont nommés « traits de personnalité ») de plus de 15000 chiens appartenant à 164 races différentes. Une partie de leur échantillon (1175 chiens) leur a servi à rechercher des traits de tempérament qui seraient caractéristiques de races (n = 47). Leurs résultats confirment une dimension individuelle des traits de tempérament au détriment d’une dimension raciale, tandis que toutes les races présentent essentiellement une tendance à jouer et à chasser. Cela pourrait traduire deux faits importants : 1° le maintien du caractère prédateur du chien même s’il ne s’exprime la plupart du temps que par une réponse de poursuite lors de la stimulation par un objet en mouvement ; 2° le caractère de dépendance et « d’infantilisation » dans lequel sont maintenus les chiens. Mais il est aussi important de se rappeler que ce que l’on appelle le jeu est un ensemble de patterns moteurs empruntés à la compétition, voire au comportement agressif et à la sexualité. Le classement de ces patterns moteurs dans un ensemble « ludique » correspond à une réelle fonctionnalité, que l’on observe par la symétrie des interactions et par la répétition de ces patterns au cours d’une succession d’interactions. Ces résultats sont en contradiction avec ce que pouvaient laisser entendre les « standards » comportementaux, en fait « tempéramentaux », établis par la FCI. Le tempérament serait donc bien d’abord une dimension génétique individuelle qui pourrait se trouver en contradiction avec les caractères recherchés en relation avec une tâche particulière utile à l’homme dans une race donnée. La difficulté d’aborder la génétique du tempérament, même s’il a par définition une forte valence génétique, réside dans la détermination du phénotype comportemental à mettre en évidence en préalable à une analyse génétique.
Vers un scénario plausible intégrant la sélection naturelle sur le tempérament
12 L’importance de la sélection de traits de tempérament est à rapprocher des scénarios de l’évolution du chien à partir de l’espèce mère, Canis lupus. Le scénario de Coppinger & Coppinger (2001) proposé plus haut postule que des loups possédant des traits de tempérament d’intrépidité seraient devenus commensaux des humains nomades, s’en approchant jusqu’à en exploiter les refus et déchets alimentaires. Cette intrépidité aurait conféré à ces loups un avantage sélectif assez évident, du fait de leur double potentialité de prédateur et de détritivore. Mais comment ce trait aurait-il pu se propager pour conduire à une radiation spécifique ? Des travaux sur deux types de populations de loups gris (Canis lupus) d’Amérique du Nord, l’une sédentaire et l’autre migrante, suggèrent comment cela a pu être possible (Musiani et al., 2007). Les loups sédentaires habitent des forêts boréales de conifères et se nourrissent de la prédation de proies de différentes tailles, mais toutes également sédentaires. La population de loups migrants vit à proximité de ces forêts dans un milieu de toundra, sans qu’il y ait de barrières géographiques, physiques ou autres entre ces populations et celles des loups sédentaires. Ces loups ont comme proie essentielle un caribou de toundra/taïga, le « Barren ground Caribou » (Rangifer tarandus groenlandicus). Cette espèce est migratrice, se reproduisant dans la toundra plus au nord des lieux fréquentés l’hiver en lisière des forêts boréales. Musiani et al. (2007) ont montré qu’effectivement une population de loups migrait en relation avec la migration des caribous alors que l’autre population était sédentaire. Ils ont montré de plus une profonde divergence génétique entre ces deux populations, à laquelle était associée une différenciation du pelage, les loups sédentaires étant plutôt sombres que pâles, tandis que les loups migrateurs sont dans leur immense majorité du type pâle. Il existerait donc une divergence génétique entre ces deux populations de loups comme conséquence de facteurs écologiques, de spécialisation du choix des proies, et en l’absence d’autres types d’obstacles au brassage génétique. Ces résultats rappellent donc le scénario du moment de la divergence avec le postulat de deux populations de loups : l’une sédentaire (la population « mère ») et l’autre migratrice suivant les mouvements de ses commensaux, les humains. On peut alors penser que le trait de tempérament « intrépide » sous fort contrôle génétique se serait fixé seulement au sein de cette dernière population sans échanges avec la population sédentaire.
Perspectives
13 Pour conclure, des arguments écologiques modernes tendraient à supporter le scénario d’une spéciation du chien à partir du loup initialement commensal de l’homme nomade. Cette sélection naturelle reposerait sur l’émergence et la propagation d’un trait de tempérament, l’intrépidité. La possession de ce trait permet à des individus de conquérir de nouvelles niches écologiques et présente donc un impact fort en termes de sélection et d’évolution. Une fois cette population de loups équipée de ce trait, une deuxième phase de sélection artificielle, où l’humain prend le contrôle de la reproduction du loup, se serait mise en place.
14 Aucun trait de tempérament ne permet actuellement de qualifier une race de chiens. Toutefois, une sélection artificielle explicite de traits adaptés pourrait permettre d’obtenir des chiens de compagnie aux morphologies diverses mais au tempérament peu varié, privilégiant par exemple la docilité ou la passivité.
15 Les traits de tempérament comme caractéristique génétique individuelle sont très vraisemblablement une expression assez directe de caractéristiques physiologiques héritées ou acquises très précocement à l’état fœtal, notamment de taux de sécrétion ou de densité de récepteurs de neuromédiateurs au niveau synaptique. L’importance de la prise en compte du tempérament dans l’évolution des espèces est attestée par le fait que quels que soient les scénarios proposés pour l’évolution du chien, à la fois Clutton-Brock (1999) et Coppinger & Coppinger (2001) mentionnent ce terme comme intervenant dans l’apparition d’espèces domestiques. Le tempérament interviendrait dans le rapprochement d’espèces sauvages et de l’homme comme une disposition individuelle, vecteur de dispositions spécifiques, dont les humains peuvent tirer ultérieurement profit et qu’ils peuvent orienter. Il serait donc extrêmement intéressant d’explorer plus en détail la relation entre traits de tempérament et génétique, et en premier lieu par une détermination précise du phénotype « tempéramental » ; en d’autres termes, catégoriser les caractères de tempérament susceptibles d’intéresser les sélectionneurs de races de chiens. Ainsi toutes les conséquences d’une sélection artificielle pourraient être explorées et maîtrisées de manière à associer ou séparer des caractères phénotypiques précis : morphologiques, de tempérament, comportementaux et de propension à développer certaines pathologies, pour une sélection de chiens mieux adaptés à un mode de vie ou à des fonctions particulières.
Extrait des standards comportementaux édictés par la Fédération cynologique internationale (FCI) tiré du site chiensderace.com/php/fiche_race.php (Anonyme, 2009). La rubrique dans laquelle sont décrits ces standards est intitulée « Caractère/Mouvement ». Beaucoup de ces standards s’apparentent à des descriptions, certains à des injonctions (Berger allemand, Boxer, etc.). [Les mentions de fonction et de caractères entre guillemets sont les transcriptions exactes du site en italiques*. NDA.]

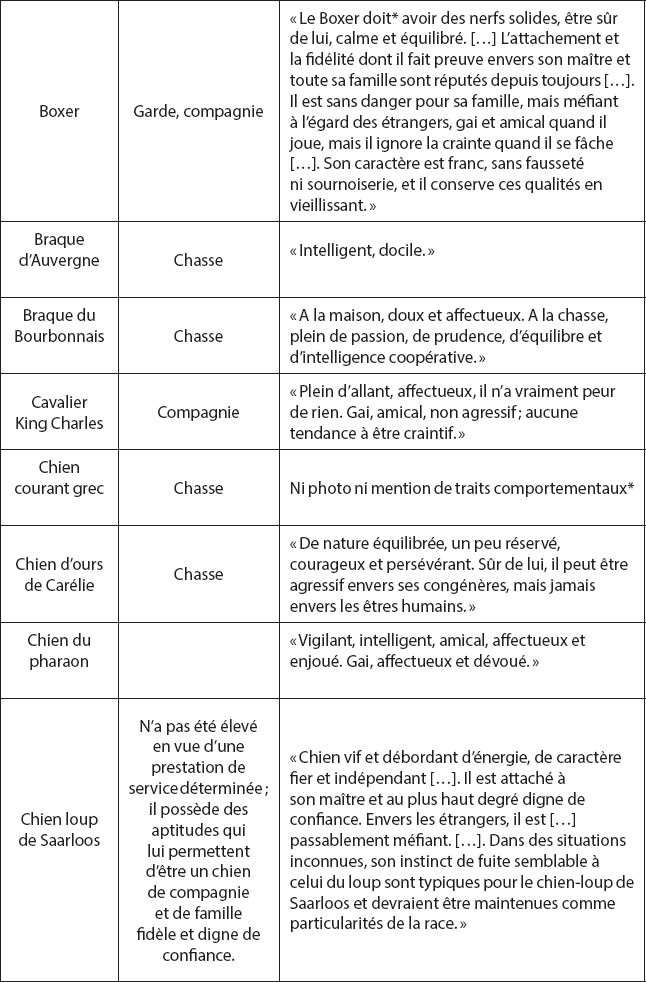


Extrait des standards comportementaux édictés par la Fédération cynologique internationale (FCI) tiré du site chiensderace.com/php/fiche_race.php (Anonyme, 2009). La rubrique dans laquelle sont décrits ces standards est intitulée « Caractère/Mouvement ». Beaucoup de ces standards s’apparentent à des descriptions, certains à des injonctions (Berger allemand, Boxer, etc.). [Les mentions de fonction et de caractères entre guillemets sont les transcriptions exactes du site en italiques*. NDA.]
Références
- Alderton D. 2002. Chiens. Bordas, Paris.
- Anonyme. 2009. chiens-derace.com : site de la Fédération cynologique internationale (FCI).
- Breton E. 2008. Spitz, chiens nordiques et chiens d’attelage : approche de leurs particularités comportementales. Thèse de doctorat vétérinaire. École nationale vétérinaire d’Alfort.
- Campan R & Scapini F. 2002. Le comportement animal : approche systémique du comportement. De Boeck, Paris.
- Clutton-Brock J. 1995. Origins of the dog : domestication and early history. In The domestic dog : its evolution, behaviour and interactions with people, pp. 7-20. J. Serpell (ed.), Oxford University Press, Oxford.
- Clutton-Brock J. 1999. A natural history of domesticated mam-mals. 2nd ed. Cambridge University Press, Cambridge.
- Coppinger R. & Coppinger L. 2001. Dogs : a new understanding of canine origin, behavior, and evolution. Univ. Chicago Press, Chicago.
- Franck H. 1980. Evolution of canine information processing under conditions of natural and artificial selection. Z. Tierpsychol. 53 : 389-399.
- Jones A.C. & Gosling S.D. 2005. Temperament and personality in dogs (Canis familiaris) : a review and evaluation of past research. Appl. Anim. Behav. Sci. 95, 1-53.
- Goldsmith H.H. 1983. Genetic influences on personality from infancy to adulthood. Child Development 54 : 331-355.
- Goldsmith H.H., Buss A., Plomin R., Rothbart M.K., Thomas A., Chess S., Hinde R.A. & Mccall R.B. 1987. What is temperament ? Four Approaches. Child Development 58, 505-529.
- Kagan J., Reznick J. & Snidman N. 1988. Biological bases for childhood shyness. Science 240 : 167-171.
- Manning A. 1972. Introduction to Animal Behaviour. E. Arnold, London.
- Musiani M., Leonard J., Cluff H.D., Gates C.C., Mariani S., Paquet P.C., Vilà C. & Wayne R.K. 2007. Differentiation of tundra/taiga and boreal coniferous forest wolves : genetics, coat colour and association with migratory caribou. Molecular Ecology 16 : 4149-4170.
- Schneirla T. 1959. An evolutionary and developmental theory of biphasic processes underlying approach and withdrawal. In Nebraska Symposium on Motivation. M. Jones (ed.). pp. 27-58. University of Nebraska Press, Lincoln.
- Savolainen P., Zhang Y-P., Luo J., Lundeberg J. & Leitnen T. (2002). Genetic evidence for an East Asian origin of domestic dogs. Science 298 : 1610-1613.
- Svartberg K. & Forkman B. 2002. Personality traits in the domestic dogs (Canis familiaris). Appl. Anim. Behav. Sci. 79 : 133-155.
- Trut L.N. 1999. Early canid domestication : the farmfox experiment. American Scientist 87 : 160-169.
- Vilà C., Seddon J. & Ellegren H. 2005. Genes of domestic mammals augmented by backcrossing with wild ancestors. Trends Genet. 21 : 214-218.
- Wayne R.K. & Ostrander E.A. 2007. Lessons multiple and ancient origins of the domestic dog. Science. 276 : 1687-1689.
- Wilson D., Clark A., Coleman K. & dearstyne T. 1994. Shyness and boldness in humans and other animals. Trends. Ecol. Evol. 9, 442-446.