Analyse cladistique ou classification cladistique ?
- Par Ernst Mayr,
- Traduit par Pascal Tassy
- et Martin S. Fischer
Pages 13 à 70
Citer cet article
- MAYR, Ernst,
- Traduit par TASSY, Pascal
- et FISCHER, Martin S.,
- Mayr, Ernst.,
- et al.
- Mayr, E.,
- Traduit par Tassy, P.
- et Fischer, M.-S.
https://doi.org/10.3917/biosy.029.0013
Citer cet article
- Mayr, E.,
- Traduit par Tassy, P.
- et Fischer, M.-S.
- Mayr, Ernst.,
- et al.
- MAYR, Ernst,
- Traduit par TASSY, Pascal
- et FISCHER, Martin S.,
https://doi.org/10.3917/biosy.029.0013
Notes
-
[1]
Les termes de cladisme et de cladistique ont été utilisés indifféremment. Comme le suffixe « -ique » correspond à celui de phénétique, de systématique et de génétique, je préfère maintenant utiliser le terme cladistique. (Note de l’auteur.)
-
[2]
Un groupe holophylétique est un groupe qui contient tous les descendants d’une espèce souche. (Note de l’auteur.)
-
[3]
En français dans le texte. (Note des traducteurs.)
-
[4]
En allemand dans le texte. (Note des traducteurs.)
-
[5]
Hennig a qualifié un groupe de paraphylétique si la ressemblance des taxons qui le composent est fondée sur la symplésiomorphie. Par exemple, les Reptilia sont un groupe paraphylétique, au contraire des archosauriens (crocodiles et oiseaux) et des thérapsides (reptiles mammaliens et mammifères) qui sont holophylétiques. (Note de l’auteur.)
-
[6]
Traduction p. 495-496 sur la dernière édition de l’Origine des espèces par Ed. Barbier. (Note des traducteurs.)
-
[7]
Traduction p. 496 sur la dernière édition de l’Origine des espèces par Ed. Barbier. (Note des traducteurs.)
J’aimerais tout particulièrement mettre en exergue que je ne veux pas discréditer un auteur au moyen d’une discussion critique. Bien au contraire, il n’est pas utile de critiquer des travaux insignifiants.
1 Le choix d’une méthode dans une discipline scientifique dépend largement des objectifs de cette discipline. Si l’on veut déterminer laquelle de plusieurs méthodes de classification de plantes et d’animaux est la plus fructueuse, on doit d’abord clarifier sa propre conception de la systématique. En conséquence, fort justement, Hennig débute Grundzüge einer Theorie der phylogenetischen Systematik (1950) par une discussion sur la systématique (p. 1-12). La systématique, dit-il, est l’action d’ordonner la diversité de la nature au moyen de la construction d’une classification qui peut servir de système de référence général. « Créer un tel système de référence général, et rechercher les relations qui en découlent en direction de tous les autres systèmes possibles de la biologie, est le but de la systématique » (Hennig 1966 : 7 [id. 1950 : 10]).
2 Le but d’un créateur de classifications, est donc de trouver le « système de référence général » le meilleur possible. Cela dit, on doit être plus explicite : une classification, à l’inverse d’un schéma d’identification, fonctionne comme une théorie biologique (avec toutes les propriétés explicatives, prédictives et heuristiques d’une théorie) (Mayr 1969 : 79-80) ; elle doit fournir une fondation sensée à toutes les études comparatives en biologie et doit être capable de servir de stockage efficace et de système de récupération de l’information (Mayr 1969 : 229-244).
3 Dans de nombreux secteurs de la biologie les généralisations sont dérivées de comparaisons. Cependant, en matière de recherches évolutionnistes, les comparaisons entre groupes ne sont significatives que si ces groupes sont correctement formés, c’est-à-dire consistent en des éléments « reliés ». La construction de classifications efficaces est donc, comme le dit Hennig, un prérequis pour de nombreux travaux en biologie. Warburton (1967) a essayé de spécifier les critères à partir desquels on peut juger qu’une classification est supérieure à une autre du point de vue des fonctions de « système de référence général », de théorie biologique et de système de stockage et récupération de l’information.
4 Ce que l’on a appelé la « nouvelle systématique » a dominé les années 1930 et 1940. Les taxinomistes ont concentré leurs efforts sur le niveau des espèces et des populations (microtaxinomie), à partir duquel le nouveau champ de la génétique des populations s’est développé. Les problèmes liés à la classification des taxons supérieurs (macrotaxinomie) furent largement négligés. Une minorité significative de chercheurs, particulièrement les paléontologues et les anatomistes comparatifs, pensait cependant qu’étaient restées sans réponse de nombreuses questions issues du credo darwinien, apparemment simple, selon lequel les classifications devaient refléter les « relations de parenté » ou « l’ascendance commune ». C’était évident en lisant plusieurs contributions à des ouvrages dirigés par Huxley (1940), Heberer (1943), Jepsen, Mayr et Simpson (1949), et plus particulièrement les écrits de Simpson (1945) et de Rensch (1947). Le ferment intellectuel de cette période aboutit dans les années 1950 et 1960 à la formulation de trois théories compétitives de classification, chacune prétendant être plus objective et être un meilleur système de référence général que les deux autres. Il s’agit donc de caractériser ces trois théories évoquées par Günther (1971, 1976) et décrites plus en détail par Mayr (1969 : 68-77). (Malheureusement, de nombreux taxinomistes n’ont pas de théorie du tout et considèrent les espèces et les taxons supérieurs de façon purement descriptive, en tenant les classifications comme de simples systèmes d’identification.)
Les trois théories concurrentes de la classification
- a. Systématique phénétique (la phénétique) : selon cette théorie, les organismes sont classifiés sur la base de la « ressemblance globale ». La ressemblance est calculée à partir de la présence ou de l’absence de nombreux caractères ou états de caractères non pondérés (Sokal & Sneath 1963). Cette méthode n’établit pas les groupes par inspection mais ordonne les unités taxinomiques inférieures (habituellement les espèces) en groupes à l’aide de procédures standardisées.
6 Les méthodes et les principes de la phénétique ont été analysés de façon critique ailleurs (Mayr 1965, 1969 : Johnson 1970 ; Hull 1970).
- b. Systématique cladistique (la cladistique) [1] : selon cette théorie les organismes sont classifiés et catégorisés exclusivement sur la base du « degré d’ancienneté de l’ascendance commune ». La parenté des espèces à l’intérieur des taxons est reconnue sur la possession commune de caractères dérivés (« apomorphes »). Le regroupement et la catégorisation sont donnés simultanément par les points de branchements. On verra plus loin pourquoi la désignation de cette théorie par l’expression « systématique phylogénétique » est trompeuse.
- c. Systématique évolutionniste : selon cette théorie les organismes sont classifiés et catégorisés sur la base de deux types de facteurs, 1) le branchement phylogénétique (« degré d’ancienneté de l’ascendance commune » défini rétrospectivement), et 2) la quantité et la nature du changement évolutif entre les points de branchement. Ce dernier facteur, en retour, dépend de l’histoire évolutive d’une branche donnée, c’est-à-dire de son entrée ou non dans une nouvelle zone adaptative et dans quelle mesure elle a éprouvé une radiation majeure. Le taxinomiste évolutionniste essaie de maximiser en même temps dans sa classification le contenu informatif des deux types de variables (1 et 2 ci-dessus).
8 La méthode de classification synthétique ou évolutionniste combine donc les composants de la cladistique et de la phénétique, mais de façon assez différente. Elle est en accord avec la cladistique à propos du postulat selon lequel la construction de la phylogénie aussi complète que possible doit précéder la classification, dans la mesure où les groupes qui ne sont pas composés de descendants d’un ancêtre commun sont artificiels et ont une valeur prédictive faible. De façon plus générale elle est aussi en accord avec la cladistique vis-à-vis de la pondération prudente des caractères. Elle rejette, cependant, le processus « divisif » de la classification (la classification « vers le bas »), qui est évident dans la définition cladistique de ce qui est « monophylétique ». La classification évolutionniste rejette la plupart des axiomes de la phénétique, mais est en accord avec elle à propos du processus de regroupement selon une approche largement phénétique. Toutefois, au contraire de l’approche non pondérée des phénéticiens, elle fonde ses conclusions sur la pondération prudente des caractères.
9 La méthode où les éléments cladistique et phénétique sont combinés a été initiée par Darwin (voir ci-dessous).
La cladistique est-elle la meilleure théorie de la classification ?
10 Les cladistes sont sincèrement convaincus que leur théorie produit la meilleure classification. Par exemple Hennig (1971 : 9) dit que « la prétention à la primauté de la systématique phylogénétique parmi toutes les formes possibles de systématique biologique n’a jamais été réfutée ne serait-ce que légèrement ». Günther (1971 : 38) dit de la même façon : « W.Hennig a élaboré et nourri sa théorie de la systématique phylogénétique au point qu’on peut la considérer comme irréfutable. » Günther (1971 : 76) affirme plus loin que parmi les trois conceptions en cours de la systématique biologique, c’est le concept phylogénétique (généalogique) cohérent qui seul permet de tirer des conclusions phylogénétiquement non équivoques. On peut trouver de telles affirmations dans les écrits de Brundin, Crowson, Nelson, Schlee et d’autres cladistes. Griffiths (1972 : 9) dit, par exemple, que la méthode de Hennig « fournit la seule base théorique sensée permettant de trouver une équivalence objective entre les taxons auxquels a été assignée une catégorie particulière dans un système phylogénétique ».
11 À partir de cette conviction de la supériorité de leur méthode les cladistes sont véritablement surpris de constater « pourquoi il y encore tant de systématiciens qui n’ont pas eux-mêmes pratiqué la systématique phylogénétique (ou seulement avec des réserves) » (Hennig 1971 : 9). Hennig répond à sa propre question en disant que l’adoption générale de la systématique phylogénétique n’est gênée que par une familiarité insuffisante avec les objectifs et les méthodes de la cladistique. Günther (1971 : 77), de son côté, pense que si la cladistique n’est pas plus généralement appliquée c’est à cause de l’ignorance ou la négligence de trois faits : 1) l’absence de caractères disponibles suffisamment distincts, 2) le manque de certitude sur l’état ancestral ou dérivé des caractères, 3) la difficulté de reconnaître clairement les convergences. En d’autres termes, les deux auteurs pensent que ce sont des raisons empiriques plutôt que conceptuelles qui sont responsables du délai dans l’adoption plus rapide de la cladistique.
12 Cette conclusion est-elle vraiment justifiée ? Est-ce qu’un arrangement purement généalogique répond à la demande d’une « meilleure classification » ? En fait, comment détermine-t-on laquelle parmi plusieurs classifications est la meilleure ?
13 Les théoriciens des classifications se sont mis d’accord depuis longtemps sur le fait que, dans la plupart des cas, les « meilleures » classifications sont celles qui permettent le plus grand nombre de conclusions et de prédictions. Mill (1874 : 466-467) l’a dit, il y a un siècle : « Les buts d’une classification scientifique sont atteints si les objets sont regroupés de telle manière que le nombre de propositions générales que l’on peut faire est le plus grand et que les propositions soient plus considérables que celles que l’on pourrait tirer de tout autre groupement des mêmes objets. »
14 Les opposants de la cladistique prétendent que les classifications cladistiques ne satisfont pas le critère de Mill de la « meilleure classification ». Le nombre d’affirmations évolutionnistes et les prédictions qui peuvent être tirées des nombreux groupes holophylétiques [2] (comme les oiseaux et les crocodiliens) sont souvent minimes, et ne consistent qu’en une liste de synapomorphies. En réalité, la théorie de la classification cladistique semble souffrir de faiblesses conceptuelles fondamentales.
15 Le débat ne peut être tranché sans chercher à analyser la théorie (en incluant toutes les implications sous-jacentes) sur laquelle est fondée la cladistique.
16 De nombreux indices laissent penser que la plupart des cladistes n’ont jamais considéré sérieusement les autres théories de la classification, particulièrement la théorie de la taxinomie évolutionniste. Pour quelle autre raison Hennig (1971 : 7) aurait classifié les taxinomistes évolutionnistes du côté de ces taxinomistes qui travaillent sans aucune théorie ? (Voir également Brundin 1972 : 111.) D’autres cladistes avancent en débattant comme si la seule alternative était entre leur théorie et la phénétique (la classification qui repose simplement sur la ressemblance). Même Griffiths (1972 : 18) qui distingue clairement les trois méthodes de classifications, n’oppose, dans sa défense de la cladistique, que des arguments envers « les classifications morphologico-phénétiques ». Les objections contre la cladistique sont balayées comme étant des incohérences ou seulement d’ordre psychologique (Günther 1971 : 38).
17 Les esprits ne se rencontreront pas si les cladistes ne font pas face aux critiques de leurs opposants et s’astreignent à les réfuter, point par point. Griffiths (1972) est le seul cladiste qui ait jamais essayé de conduire une telle réfutation.
18 À l’inverse du flot de publications en faveur de la cladistique de ces dernières années (par Bigelow, Brundin, Cracraft, Crowson, Griffiths, Günther, Hennig, Kiriakoff, Nelson, Rosen, Schlee et autres) on compte peu d’analyses critiques de cette théorie. Simpson (1961) et moi-même (Mayr 1969) l’on fait en passant [3] dans des manuels importants. Il y a eu plusieurs courtes critiques d’ouvrages ; quelques points spécifiques (comme la définition de la « monophylie ») ont été critiqués par Ashlock, Colless, Farris, Gutmann, Johnson, Michener, Peters, et d’autres. Mais les articles de Darlington représentent le seul essai sérieux de critique à grande échelle de la cladistique. Et encore, cette critique était plus dirigée vers l’application de la cladistique à la biogéographie qu’aux principes de base de la théorie. En fait, Darlington (1970 : 1) lui-même a signifié que sa critique « n’était pas une considération générale de la cladistique ». L’objectif de la présente analyse critique est de combler ce sérieux hiatus dans la littérature taxinomique.
Les composants de la cladistique
19 Pour bien comprendre la cladistique il est important de réaliser que celle-ci consiste en réalité en deux ensembles d’opérations différents : 1. la reconstruction du schéma de branchement de la phylogénie au moyen de l’analyse cladistique, 2. la construction de la classification cladistique fondée sur ce schéma de branchement.
20 La première de ces deux opérations est importante et ne souffre guère d’objection. C’est la seconde qui a été largement critiquée et qui sera soigneusement analysée dans les pages suivantes, avec une attention particulière à la prétention cladiste qu’une classification doit être l’image du schéma de branchement de la phylogénie.
La reconstruction du schéma de branchement de la phylogénie (analyse cladistique)
21 L’analyse cladistique part de l’idée fondamentale qu’une classification sensée ne peut être construite sans une compréhension précise de la phylogénie d’un groupe donné. Les taxinomistes évolutionnistes sont d’accord, en gros, avec cette idée. Toute phylogénie, en dehors des cas d’évolution réticulée, est strictement généalogique. Hennig a tout à fait raison quand il écrit : « La recherche phylogénétique en tant que science biologique est concevable uniquement si l’on admet que le premier objectif est la découverte de la relation généalogique entre les espèces » (Hennig 1969 : 33).
22 Mais comment procéder si l’on veut reconstruire la phylogénie d’un groupe ? Comme Hennig (1969 : 19) l’a dit, la méthode repose en dernière analyse sur le fait que « toutes les différences et ressemblances entre diverses espèces sont le fruit de la phylogénie. Durant sa subdivision, une espèce transmet ses caractères à ses espèces filles qu’ils soient transformés ou non transformés ». Hennig est tout à fait conscient que tout ce que l’on peut inférer à l’aide de cette méthode est la séquence des subdivisions mais pas la chronologie absolue.
23 Hennig a souligné, et à juste titre, qu’une phylogénie n’a pas besoin d’être fondée sur des fossiles mais peut être inférée à l’aide d’une analyse comparative attentive des caractères morphologiques. Cette thèse est bien illustrée par la classification des mammifères actuels. Nos idées sur leurs relations de parenté, fondées sur l’anatomie comparée, n’ont en rien été réfutées par des découvertes ultérieures dans les archives fossiles. En revanche, les archives fossiles sont tout à fait indispensables pour la détermination des chronologies absolues.
24 L’étape la plus importante de l’analyse cladistique est l’essai de séparer les caractères en caractères ancestraux (plésiomorphes) et dérivés (apomorphes). Seuls les derniers sont tenus pour une preuve légitime de relations de parenté, et, en conséquence, les taxons sont fondés par la possession commune de caractères dérivés (synapomorphies). (Pour une considération de la valeur des symplésiomorphies dans le processus de catégorisation, voir page 57.) Hennig et aucun de ses successeurs n’a jamais prétendu que ce principe important était nouveau. « Faire observer que les taxons ne doivent, dans leur groundplan, être caractérisés que par des états apomorphes (dérivés), n’est, bien sûr, pas nouveau et semble pour le plus grand nombre, aller de soi » (Griffiths 1972 : 21). Un seul exemple : c’est sur ce principe que la classification des Perlaria par Tillyard (1921 : 35-43) était fondée. En fait, on peut dire que la plupart des meilleurs taxinomistes des époques anciennes ont appliqué ce principe, comme on peut le constater en étudiant leurs classifications.
25 Néanmoins on doit porter crédit à Hennig d’avoir entièrement développé les principes de l’analyse cladistique. La reconnaissance claire de l’importance des synapomorphies pour la reconstruction de la séquence des branchements est la contribution majeure de Hennig. Les cladogrammes qui sont construits à l’aide de cette méthode sont aussi importants pour le taxinomiste évolutionniste qu’ils le sont pour le cladiste. J’ai déjà attiré l’attention (Mayr 1969 : 212-217) sur la valeur extrême de cette méthode pour délimiter les taxons. La séquence temporelle relative des différents points de branchement que fournit le cladogramme est de grande valeur pour de nombreuses études, particulièrement en zoogéographie, comme Hennig (1960) lui-même l’a illustré à propos des diptères de Nouvelle-Zélande.
Analyse cladistique et classification cladistique
26 Il n’y a guère d’opposition entre taxinomistes cladistes et évolutionnistes à propos des cladogrammes qui résultent de l’analyse cladistique. L’opposition vient de la relation entre le cladogramme et la classification qui en est issue. Les cladistes affirment qu’il existe une relation univoque entre le cladogramme (schéma phylétique) et la classification. Une fois, construit, le cladogramme produit automatiquement, pourrait-on dire, la classification. Le cladogramme et la classification sont pour les cladistes les deux faces de la même pièce. Le taxinomiste évolutionniste, au contraire, croit qu’un simple schéma de branchement ne peut pas donner une information aussi intéressante qu’une classification évolutionniste qui, elle, prend en considération d’autres processus évolutifs (voir ci-dessous).
27 Traditionnellement, la première étape suivie par les taxinomistes praticiens en matière de classification des animaux a toujours été la délimitation de groupes apparemment « naturels » en les examinant. Au départ, ces derniers sont clairement fondés sur la « ressemblance » apparente, c’est-à-dire un critère phénétique. Quand Hennig (1950) a présenté pour la première fois la méthode cladistique, pratiquement tous les taxons supérieurs d’animaux étaient déjà connus. Il a donc en conséquence adopté automatiquement la méthode taxinomique traditionnelle de catégorisation et de regroupement de taxons animaux que d’autres auteurs avaient délimité préalablement. La validité de ces groupes provisoires est ensuite testée en taxinomie traditionnelle au moyen d’une série de critères additionnels, comme l’homologie des caractères (ressemblants et non ressemblants), la présence de synapomorphies, la relation chronologique à des groupes similaires, l’absence de conflit avec les archives fossiles (s’il y en a), l’absence de convergence (= fausse ressemblance), une distribution géographique qui a du sens, etc. Plus le taxinomiste est expérimenté, plus il peut entreprendre ces tests rapidement et profondément. (Le procédé de classification du phénéticien est totalement différent). Une fois établie, la classification est alors régulièrement améliorée par le procédé appelé par Hennig (1950) « illumination réciproque », dont Hull (1967) a montré qu’il ne tenait pas du raisonnement circulaire. En fait, la méthode n’est rien de plus qu’une autre application de l’approche hypothético-déductive (Popper 1959, 1963) si communément appliquée dans toutes les branches de la science et particulièrement (depuis Darwin) en biologie. Dès qu’une classification nouvelle (ou révisée) est proposée, elle engendre de nouvelles informations qui, en retour, entraîneront une nouvelle analyse et une possible amélioration de la classification. Cette approche traditionnelle de la classification a été suivie pendant le 19e et le 20e siècle sans critique sérieuse.
28 Malheureusement cette approche par essai et erreur est quelquefois inefficace et a entraîné de fréquents changements de classifications. L’espoir a donc été mis, encore et toujours, dans de meilleures approches. Les meilleurs taxinomistes sont d’accord sur deux points minimaux, l’importance cruciale du bon choix des caractères (déjà soulignée par Darwin), et la nécessité de fonder les taxons sur de nombreux caractères. Mais même cela ne réduit pas totalement les incertitudes.
29 C’est la proposition nouvelle de Hennig de transcrire simplement le cladogramme en une classification hiérarchique et de se libérer ainsi de toutes ces incertitudes. Toutefois, construire une classification directement à partir du cladogramme n’est pas une proposition qui intègre la totalité de la théorie de la classification cladistique. Pour cette raison il serait hautement désirable de présenter une présentation détaillée de la théorie entière et de toutes ses ramifications, mais ce serait assez difficile. Non seulement parce que le travail original de Hennig (1950) est écrit dans un style obscur, mais quelques-unes de ses anciennes thèses n’apparaissent plus dans ses écrits récents. En plus, il semble que certains de ses successeurs, comme Brundin, Schlee et Griffiths, ont ajouté des postulats qui, s’ils étaient peut-être implicites dans la théorie originale, n’ont pas été présentés explicitement par Hennig lui-même. Toutefois, les principaux postulats et les principales thèses ont été signifiés suffisamment souvent pour qu’on puisse les énumérer. J’essaierai de faire la liste des plus importants, de préférence en citant directement les travaux des cladistes. Je suppose que ma liste n’est pas complète ni que les postulats ont été adoptés par tous les cladistes. Cela dit, j’espère que cette liste servira de base convenable pour toute analyse ultérieure.
30 Les points suivants sont les postulats les plus importants de la cladistique (les pages données ci-dessous entre parenthèses font référence à la discussion plus détaillée qui suit) :
- tous les taxons doivent être « monophylétiques », ce terme ayant été redéfini de façon nouvelle, en conflit avec sa définition traditionnelle (p. 31) ;
- le terme phylogénétique doit être appliqué, de façon restreinte, au branchement dans la phylogénie (p. 26) ;
- la relation de parenté est mesurée en termes de « degré d’ancienneté de l’ascendance commune », c’est-à-dire étroitement généalogique (p. 27-31) ;
- il y a « une seule dimension dans la phylogénie et c’est la dimension du temps » (Brundin 1966). Par voie de conséquence, la subdivision des lignées phylétiques (reconstruites à partir de l’acquisition conjointe de caractères dérivés) est tenue pour la seule preuve légitime de la construction de la classification. Prendre en compte également les ressemblances ou la quantité relative de caractères ancestraux (plésiomorphes) aboutirait à un « système syncrétique » qui « ôterait à la combinaison toute valeur scientifique » (Hennig 1966 : 77). Ce dernier cite en l’approuvant l’affirmation de Bigelow (1956) selon laquelle « la classification doit être fondée sur l’une ou l’autre (“ressemblance globale” ou “degré d’ancienneté de l’ascendance commune”…), pas sur les deux, si l’on veut éviter toute confusion philosophique » (Hennig 1966 : 77) (p. 28, p. 64) ;
- la catégorie à laquelle est assigné un taxon est donnée automatiquement par l’âge géologique absolu de l’espèce souche ou (selon une formulation moins rigoureuse) par « l’âge relatif » de l’espèce souche (p. 49). (Voir également Crowson : 251 et critique par Griffiths [1972 : 10, 16].) ;
- les espèces peuvent être délimitées dans le temps par deux événements successifs de spéciation (p. 40) ;
- le processus de subdivision des lignées est toujours une dichotomie, qui résulte dans la production de deux espèces sœurs (p. 40) ;
- « l’homologie […] est habituellement définie en termes d’origine commune dans le temps » (Griffiths 1972 : 17). (Ce n’est tout simplement pas vrai. À l’exception d’une relation directe d’ancêtre à descendant, le concept d’homologie est totalement indépendant de la dimension temporelle. Aucun autre cladiste n’a avancé une telle affirmation. Hennig lui-même a adopté le concept d’homologie de Remane 1952.) ;
- toute classification est « fondamentalement » une classification horizontale, uniquement valable pour une période de temps donnée (Hennig 1950 : 259) et, par conséquent le même taxon doit être assigné à des catégories différentes dans les différentes périodes géologiques (p. 49-50).
Objections à l’encontre de la théorie de la classification cladistique
32 Le postulat de base de la théorie cladistique, une congruence absolue entre le cladogramme et la classification, ne peut être satisfait qu’en faisant de nombreuses hypothèses et redéfinitions et en ignorant de nombreux faits de l’évolution et de la phylogénie (au sens large). Il en résulte de nombreux raccourcis théoriques et pratiques qui sont maintenant examinés, point par point, dans trois sections principales et quelques autres secondaires.
1. Décisions arbitraires
33 Afin que leur méthode de classification soit fonctionnelle, les cladistes ont pris une série de décisions arbitraires qui comprennent la redéfinition de nombreux termes, une réinterprétation de l’évolution adaptative et la proposition d’une nouvelle définition de l’espèce. Si l’on rejette ces décisions arbitraires, il ne reste plus beaucoup de raisons de classifier de façon cladistique.
a. Redéfinition de termes bien connus
34 La controverse entre les cladistes et leurs opposants est due pour une large part au fait que les cladistes ont donné un sens entièrement nouveau à des termes évolutionnistes qui ont été largement employés avec un sens différent et de façon cohérente depuis près de 100 ans. Le transfert de termes bien connus et universellement compris vers de nouveaux concepts ne peut pas ne pas entraîner de confusion. C’est particulièrement vrai pour trois termes : phylogénie, relations de parenté et monophylétique.
Phylogénie
35 Depuis l’époque de Darwin et de Haeckel le terme de phylogénie a été appliqué à tous les aspects de la descendance. Tout article ou livre des cent dernières années a utilisé le mot phylogénie (et son adjectif phylogénétique) pour tous les phénomènes révélés par la Stammbaumforschung [4]. Le Collegiate Dictionary of Zoology, par exemple donne la définition : « 1. Les relations évolutives et lignes de descendance de tout taxon. 2. L’origine et l’évolution des catégories taxinomiques supérieures » (Pennak 1971 : 395). Mais Hennig essaie maintenant de restreindre le terme à un seul aspect de la phylogénie, le branchement. Il dit : « Nous appellerons “relations phylogénétiques” les […] relations (généalogiques) entre différentes sections (dans le schéma), chacune borné par deux processus de clivage dans la séquence des individus qui sont connectés par des relations tokogénétiques » (Hennig 1966 : 20). Ou : « Nous avons défini les relations phylogénétiques […] comme ces segments du cours des relations généalogiques qui se situent entre deux processus de spéciation » (Hennig 1966 : 29). Schlee (1971) définit la phylogénie comme « l’origine des taxons, c’est-à-dire cette partie de l’évolution qu’on a désignée comme sa composante cladistique ».
36 La définition et l’usage du terme phylogénétique par Hennig sont clairement en contradiction avec l’usage universel du terme jusqu’alors. Ses schémas sont des cladogrammes et pas des arbres phylogénétiques qui par la longueur de ses branches et l’angle qu’elles font entre elles donnent bien plus d’informations qu’un cladogramme. Si la théorie spécialisée de la classification selon Hennig continuait être dénommée classification « phylogénétique » cela ne ferait qu’aggraver la confusion.
37 Gisin (1964) a proposé de qualifier la méthode de Hennig par l’expression classification « généalogique », dans la mesure où les relations généalogiques (lien de parenté) sont un aspect important de la méthode de Hennig. Malheureusement, la dénomination « généalogique » ne différencie pas la cladistique de certains autres types de classifications, car toute classification dont les taxons consistent en espèces qui descendent d’un ancêtre commun est une classification généalogique. Darwin, de la sorte, avait raison de dire que les classifications évolutionnistes sont des généalogies. Dans toute phylogénie sans hybridation (et c’est le cas de presque toutes les phylogénies animales) il ne peut exister qu’une seule généalogie, parce que chaque événement de spéciation est unique. Le branchement est une composante non équivoque de la phylogénie, ainsi que souligné par les cladistes. Toutefois même une généalogie non équivoque peut être transcrite sous la forme de plusieurs classifications. Et c’est là que se séparent évolutionnistes et cladistes. Celui qui pondère de différentes façons différents processus et événements adaptatifs (par exemple un plus grand poids appliqué à l’occupation de zones adaptatives majeures) peut aboutir à une classification très différente de celle obtenue par quelqu’un qui utilise le branchement comme seul critère (comme font les cladistes), même si tous les deux se fondent sur la même généalogie. Comme le terme « généalogique » ne peut discriminer les classifications évolutionniste et cladistique, je préfère le terme « cladistique » pour la méthode de Hennig car il s’applique à elle sans la moindre équivoque tout en ne s’appliquant à aucun autre système de classification. De plus cela correspond à la terminologie proposée par Rensch (1947) et Cain & Harrison (1960). C’est le seul terme qui introduit de façon adéquate l’insistance de la méthode de Hennig sur le branchement, et le branchement seul.
38 Quand on l’emploie à propos de phylogénie, le terme « généalogique » est évidemment utilisé sans un sens général, où les « taxons » correspondent aux « individus » d’une généalogie conventionnelle. La seule autre possibilité serait de classifier des « générations » d’individus. De fait, cela serait logiquement impeccable mais inutile pour les besoins d’une classification biologique.
Relation de parenté
39 L’expression relation de parenté a été utilisée de nombreuses façons. Le recours à un dictionnaire ne sert à rien car il mentionne des définitions extrêmement divergentes. L’expression relation de parenté (ou affinité) était très commune dans la littérature taxinomique du 18e siècle, bien avant que l’on adopte la théorie de l’évolution. Pour la plupart des auteurs de cette époque cela signifiait simplement la ressemblance. Cela dit, même aujourd’hui, relation de parenté est définie de différentes façons.
40 Les phénéticiens, au début, procédaient sur la base de l’hypothèse selon laquelle le phénotype est un reflet adéquat du génotype et que la détermination non pondérée de la « ressemblance globale » permettait une détermination correcte de la relation de parenté. Ils ne sont plus aussi dogmatiques qu’en 1963, mais pour eux relation de parenté signifie toujours ressemblance. La restriction de cette interprétation a été soulignée par Mayr (1969) et bien d’autres.
41 Les cladistes vont vers un autre extrême en restreignant le terme relation de parenté au sens de lien dans un contexte strictement généalogique. Selon Hennig (1966 : 74), la « mesure de la relation de parenté au sens phylogénétique est le degré relatif d’ascendance commune. La relation de parenté entre deux espèces est mesurée par le nombre de points de branchements qui les sépare de l’ancêtre commun » (Hennig 1950 : 129). Quand Cracraft (1972 : 381) prétend que la méthode cladistique « est la meilleure pour déterminer les relations de parenté d’une façon relativement non ambiguë », il est victime d’un raisonnement circulaire car il utilise la définition cladistique hautement spécialisée de la notion de relation de parenté. Répétons-le, la relation de parenté des cladistes est le lien généalogique. Mais pour un évolutionniste, le lien cladistique seul n’est qu’un aspect de la documentation de la relation de parenté parce qu’il ignore le destin des lignes phylétiques après le clivage.
42 Je m’explique. Comme une personne a reçu de son père la moitié de ses chromosomes et que son fils reçoit à son tour de lui la moitié de ses chromosomes, on peut dire que cette personne est génétiquement aussi étroitement liée à son père qu’à son fils. Lorsqu’il s’agit de parents collatéraux (jumeaux, cousins) et de descendants plus distants (grands-parents et petits enfants, etc.), le pourcentage des gènes partagés (relations génétiques) devient presque imprévisible en raison du côté aléatoire des crossing-over et de la distribution au hasard des chromosomes homologues durant la méiose. Deux cousins germains (et même deux frères dans ce cas) peuvent entre eux avoir cent fois plus de gènes en commun que par rapport à un troisième cousin germain (ou frère) (parmi les loci variables dans cette population). Plus le nombre de générations impliquées est grand, plus grande sera la différence entre les liens généalogiques et la ressemblance des génotypes, même si tous ces parents ont dérivé leurs gènes du pool génétique d’une seule espèce.
43 En matière de phylogénie, où des milliers et des millions de générations sont impliqués, c’est-à-dire des milliers et des millions d’occasions de changement dans les fréquences de gènes, à cause des processus stochastiques, la recombinaison, la sélection et la révolution génétique, il devient absolument sans signification d’exprimer les relations de parenté uniquement en termes de liens généalogiques.
44 En plus des processus cytogénétiques, de nombreux aspects de la sélection peuvent entraîner un degré de changement génétique très distinct dans les différentes lignes de descendance. L’une des nombreuses lignes phylétiques sœurs peut pénétrer dans une nouvelle zone adaptative et être exposée à de sévères nouvelles pressions de sélection. Le résultat est que cette ligne divergera de façon spectaculaire de son proche parent cladistique et pourra tellement s’éloigner génétiquement qu’il serait biologiquement trompeur de continuer de qualifier ces groupes frères de proches parents. Cela dit, comme ils sont les descendants d’une espèce souche on doit les désigner sous le terme de groupes frères. Et en tant que groupes frères on doit les coordonner au niveau de la catégorie, c’est-à-dire selon la théorie cladistique, leur attribuer le même rang dans la hiérarchie (Hennig 1966 : 139). Cette décision ignore le fait que l’un est resté très semblable à l’espèce souche tandis que l’autre a évolué dans un type d’organisme remarquablement différent.
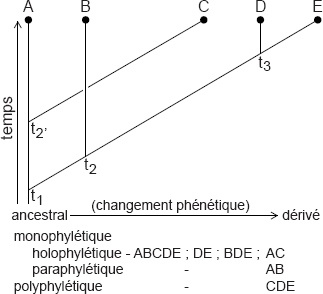
45 Cette situation est mieux illustrée par un schéma (Fig. 1). Il existe une différence génétique maximale de 25 % entre les génomes B et C, mais de 60 à 70 % entre C et D. Le cladiste dira que C est plus apparenté à D qu’à B, l’évolutionniste et le phénéticien diront que C est plus proche de B que de D.
Pourcentage inféré de différences à partir de l’ancêtre (A). Le taxon C est plus étroitement lié à B qu’à D même s’il partage un ancêtre commun plus récent avec D.

Pourcentage inféré de différences à partir de l’ancêtre (A). Le taxon C est plus étroitement lié à B qu’à D même s’il partage un ancêtre commun plus récent avec D.
46 L’indépendance des dérives adaptatives vis-à-vis du clivage phylétique est la raison pour laquelle le taxinomiste évolutionniste adopte une définition très différente de la relation de parenté. Pour lui la relation de parenté signifie la quantité partagée de génotype supposée, elle signifie le contenu génétique plus que le lien généalogique formel. Ce qui est d’intérêt primordial pour un taxon, son rôle évolutif, son système d’adaptations, et toutes les corrélations entre sa structure et ses caractères est, en fin de compte, encodé dans son génotype.
47 Comme des parents distants ne peuvent pas être analysés génétiquement, et comme une pure addition de systèmes adaptatifs complexes serait de toute façon sans raison, il est nécessaire d’inférer le degré de relation de parenté génétique sur la base de données indirectes. Selon cette vue, on utilisera toute preuve accessible, mais surtout toute combinaison de branchement phylétique et d’analyse phylétique soigneusement pondérée. Le taxinomiste évolutionniste pense qu’une approche qui superpose une analyse phénétique soigneusement pondérée sur une analyse cladistique préalable est à mieux d’établir le degré de parenté qu’une pure approche cladistique ou qu’une approche phénétique non pondérée. Une classification fondée sur une telle définition multiple des relations de parenté sera plus fiable et plus prédictive qu’une classification fondée sur des critères tirés d’un seul type d’approche. Les critères qui sont employés pour effectuer une telle pondération et la rendre significative ont été discutés ailleurs (Mayr 1969 : 217-228). Ces critères permettent de mesurer quelque chose qui est plus qu’une « ressemblance globale ».
En raison de l’évolution parallèle, le taxon C ressemble plus à D et à E qu’à A avec qui il partage un ancêtre commun proche. L’inverse est vrai pour B (d’après Ashlock 1969).
En raison de l’évolution parallèle, le taxon C ressemble plus à D et à E qu’à A avec qui il partage un ancêtre commun proche. L’inverse est vrai pour B (d’après Ashlock 1969).
48 La convergence, le parallélisme et l’évolution en mosaïque sont des phénomènes de l’évolution qui soulignent l’importance du génotype invisible sous-jacent. Un schéma (Fig. 2) illustre les difficultés créées par le potentiel dissimulé du génotype. Si l’espèce C en raison de son génotype caché acquiert en parallèle à peu près les mêmes caractères que D et E, même si elle est branchée sur la ligne qui mène à A, il serait légitime de la classifier avec D + E. Le cladiste qualifierait vraisemblablement de polyphylétique le taxon composé de C + D + E comme si la ressemblance était due à la convergence. Du point de vue du phénotype, de fait, C + D + E est un groupe polyphylétique. De nombreux phénomènes évolutifs indiquent l’existence d’un génotype caché que l’on ne peut pas tirer entièrement et directement du phénotype visible. La plupart de nos difficultés avec la polyphylie apparente sont dues aux manifestations de ce génotype caché.
Monophylétique
49 Le procédé traditionnel de reconnaissance d’un taxon supérieur est « l’intuition », c’est-à-dire une reconnaissance sur la base de caractéristiques partagées, du rôle évolutif, etc. Comme Hennig (1966 : 146) l’a écrit : « En taxinomie le but de regrouper peut démarrer en supposant que le degré de ressemblance entre les espèces correspond au degré des liens phylogénétiques, une supposition qui doit être testée soigneusement ultérieurement. » Il y a donc un accord entre Hennig et les taxinomistes traditionnels sur le fait que la caractérisation et la délimitation des groupes ont la primauté sur l’opération classificatoire (voir ci-dessus p. 22). Cette primauté est confirmée par Griffiths (1972 : 7) qui propose « que le système phylogénétique soit issu d’une révision du système traditionnel linnéen plutôt que d’une classification à part ».
50 Ce n’est qu’après avoir établi provisoirement un taxon quelconque que le taxinomiste se demande si ledit taxon est « naturel », c’est-à-dire s’il est composé d’espèces qui sont les plus proches parentes. Une façon de remplir cette condition est de se demander si tous les membres du taxon sont les descendants d’un ancêtre commun, autrement dit si le taxon est monophylétique. Le terme monophylétique est un adjectif qualificatif qui s’applique à un nom, celui de groupe (ou taxon). Parmi tous les groupes (et taxons) possibles il y en a certains que nous pouvons identifier comme monophylétiques. C’est un terme clairement « rétrospectif » (Mayr 1969 : 75). Pour cette raison, le terme monophylétique, même depuis Haeckel, a été appliqué aux groupes qui satisfont deux conditions : 1) on pense qu’en raison de leurs caractéristiques les espèces qui composent le groupe sont les plus proches parentes les unes des autres, et 2) on infère que toutes sont descendues du même ancêtre commun. Le deuxième critère est nécessaire si l’on veut exclure des groupements non naturels dus à la convergence.
51 Quand on applique aux taxons cette définition traditionnelle de monophylétique, on rencontre rarement des difficultés : les oiseaux sont monophylétiques, les crocodiles sont monophylétiques, et les reptiles sont monophylétiques. Le concept est entièrement dépourvu d’ambiguïté, même si son application rencontre parfois des difficultés, comme dans le cas de la transition thérapsides-mammifères (Simpson 1961 : 124-125 ; mais voir Crompton & Jenkins 1973).
52 Hennig a créé une confusion énorme en ajoutant à la définition traditionnelle du taxon monophylétique la qualification supplémentaire : « […] et qui inclut toutes les espèces qui descendent de l’espèce souche ». Cette définition est la conséquence inévitable de l’élimination par le concept de phylogénie selon Hennig de toute considération sur l’évolution adaptative. Puisque les oiseaux et les crocodiles (à l’exclusion de tout autre reptiles vivants) sont dérivés d’un ancêtre commun sa méthode force les cladistes à reconnaître un taxon pour l’ensemble des oiseaux et crocodiles, même si c’est un regroupement sans utilité. Hennig a transféré au mode de descendance le qualificatif « monophylétique » qui s’applique initialement au taxon. Il a fait d’un principe rétrospectif un critère prospectif. D’où l’ignorance, et même la dissimulation délibérée, des aspects le plus intéressants de l’évolution et de la phylogénie, ceux liés à la radiation adaptative et l’invasion de nouvelles zones adaptatives (discutées plus loin dans la section « Grades »).
53 Sa définition de monophylétique force Hennig à créer un nouveau terme qui s’ajoute à la terminologie phylogénétique : paraphylétique [5]. Un groupe taxinomique est paraphylétique s’il a donné naissance à des lignes collatérales spécialisées qui ne sont pas considérées comme faisant partie du groupe. Par exemple, les Reptilia sont – pour Hennig – un groupe paraphylétique, parce que des membres des Reptilia sont les espèces mères d’autres Reptilia d’une part et des oiseaux d’autre part (même chose pour l’espèce souche qui a donné naissance à la branche mammalienne). Désigner ces groupes comme paraphylétiques m’apparaît comme une approche purement formelle. De toute façon, cela n’est d’aucune signification pour les relations entre les crocodiliens et les autres reptiles de savoir que la branche qui mène aux crocodiliens (la lignée archosaurienne) a produit une branche divergente qui est finalement devenue la classe Aves. Les taxinomistes des animaux ne classifient pas des schémas de logiciens, mais, en réalité, des groupes concrets d’organismes. Notre jugement sur la classification biologique des crocodiliens n’est en rien influencé par le fait de savoir ou non qu’une branche latérale a donné naissance à un groupe fils extrêmement modifié.
54 Prendre le terme traditionnel monophylétique et le transférer vers un nouveau concept pour lequel il n’a jamais auparavant été utilisé, est contraire aux pratiques linguistiques sensées et aux principes de la terminologie scientifique. Quand les disciples de Hennig critiquent leurs opposants pour leur « usage illogique du mot monophylétique », je trouve cela grotesque. Si l’on veut un mot pour « l’agrégation de tous les groupes qui descendent d’un ancêtre commun », on peut concevoir un nouveau terme. Ashlock (1971 : 65 ; 1972) a parfaitement vu cela et a proposé le terme « holophylétique ». Ce dernier correspond à « monophylétique » tel qu’il est utilisé par Hennig à l’encontre de l’usage traditionnel.
55 Aucun cladiste ne semble s’être aperçu de quelques-unes des conséquences de la redéfinition du terme monophylétique. En étant forcé d’abandonner la classification vers le haut, comme le taxinomiste empirique l’a pratiqué depuis Darwin et même avant, on la remplace par la classification « vers le bas ». Bien qu’à partir de prémisses entièrement différents, sur un plan méthodologique le cladiste est retourné à la méthode « divisive » de la classification qui a dominé depuis Cesalpino jusqu’à Linnaeus. Son critère de division est évidemment fort différent de celui des adhérents de la logique de division d’Aristote, mais les principes de classification des deux écoles (cladistes et logiciens) sont très semblables.
56 Cela peut paraître une platitude de dire que lorsque l’on classifie, on a à traiter les entités qu’on a en face de soi. Le phénéticien et l’évolutionniste classent les espèces et les genres de cette manière. Pas le cladiste qui traite les quantités inconnues produites par les clivages phylogénétiques. De façon implicite, ses principes le forcent à prédire que les lignes sœurs qui dérivent d’une même espèce souche auront des devenirs évolutifs suffisamment similaires pour que les groupes frères qui en résultent puissent être catégorisés au même niveau (= sont coordonnés). Le cas des oiseaux et des crocodiles en est une illustration particulièrement convaincante parmi les milliers de situations où cette prédiction ne se réalise pas. C’est l’abandon du principe de classification vers le haut, qui domine depuis Darwin, et son remplacement par la classification vers le bas d’Aristote qui est l’erreur fatale de la philosophie de la classification cladistique.
57 Ceux qui adopteraient les trois termes de phylogénie, relation de parenté, et monophylétique dans leurs nouveaux sens hennigiens aberrants seraient forcés, d’assumer des changements considérables de la théorie tout entière de la phylogénie et de la classification, et de sa pratique.
58 Il est tout à fait vrai que l’on peut opérer en toute logique dans le cadre de ces nouvelles définitions. Cela dit, comme Ghiselin l’a souligné si souvent, on peut opérer de façon entièrement logique sur la base de prémisses totalement fausses. De nombreuses prétentions des cladistes, sinon la plupart, sont la conséquence de leurs nouvelles définitions de ces trois termes.
b. La négligence de la double nature du changement évolutif
59 Darwin a clairement vu que la spéciation implique deux processus indépendants. L’un est l’acquisition de l’isolement reproductif, un prérequis qui prévient de l’hybridation entre les deux espèces naissantes. L’autre est l’intégration de niches différentes d’où résulte « la divergence des caractères » dans le but de surmonter les effets de la compétition (Darwin 1859 : 111).
60 Ce qui est vrai au niveau des espèces l’est également en macroévolution. Nous pouvons distinguer deux processus de l’évolution, celui du clivage des lignes phylétiques et celui de l’invasion par les lignes phylétiques de nouvelles niches adaptatives et de zones adaptatives majeures. Toute théorie de la classification qui ne fait pas attention à l’extraordinaire différence entre les dérives des lignes phylétiques en direction de niches mineures et en direction des zones adaptatives entièrement nouvelles, est condamnée à produire des classifications qui ne sont pas équilibrées et sont sans signification. Mais cette négligence des différents aspects de l’évolution phylétique est précisément ce qu’exige la méthode cladistique.
61 Le cladiste procède dans sa construction de la classification comme si le clivage des lignées était le seul processus phylogénétique et comme si ces clivages étaient tous équivalents. Tous les clivages ont le même poids pour les cladistes, de même que les caractères ont tous le même poids pour les phénéticiens. Qu’il soit uniquement concerné par le clivage est confirmé par Hennig en maintes occasions mais ne sera exprimé ici que par une citation : « Les processus de clivage d’une espèce sont la propriété caractéristique de l’évolution ; ce fait est un point décisif. Ce sont les seuls processus historiques que l’on peut démontrer positivement, et qui ont lieu dans la nature dans les groupes supra-individuels » (Hennig 1966 : 235). Cela entraîne Hennig à prétendre que sa méthode est la seule qui donne des réponses historiques correctes.
62 Le cladiste affirme clairement que le branchement est le seul aspect de la phylogénie qui l’intéresse. Il considère comme sans importance que certaines lignées qui résultent du branchement pénètrent dans de nouvelles zones adaptatives et deviennent alors extraordinairement différentes des lignées plus conservatrices ; si elles sont issues de l’ancêtre commun par le même nombre de spéciations, on leur assignera le même rang taxinomique. Les cladistes traitent tous les caractères apomorphes de la même manière. Les taxons qui sont combinés sont ceux qui ont le plus grand nombre d’apomorphies en commun (comme étant dérivés de la même espèce souche). D’après ce que je peux en juger selon ma lecture de la littérature cladistique, les apomorphies ne sont pas pondérées, et les caractères dérivés qui sont liés à la pénétration dans de nouvelles zones adaptatives (comme dans le cas des oiseaux) n’ont pas plus de poids que les apomorphies partagées par les oiseaux et les crocodiles (qui distinguent ces archosauriens vivants des autres reptiles vivants). La même pondération est appliquée aux spécialisations mineures et aux innovations adaptatives majeures. Que certains événements de l’évolution adaptative soient plus importants que d’autres est complètement ignoré. C’est, probablement, la différence la plus significative entre les classifications cladistique et évolutionniste.
63 Il est évident que les cladistes font preuve d’une grande ambivalence dans le traitement de la divergence. Ils reconnaissent en passant le fait qu’il y a des différences dans la vitesse d’évolution des communautés de descendance mais ne tirent aucune conclusion de cette observation. Quand ils construisent leur classification, Hennig et Brundin, et aucun de leurs disciples (par exemple Schlee, Nelson, Griffiths), ne portent la moindre attention à ces différentes vitesses mentionnées par Hennig. De la même façon ils ignorent d’autres phénomènes évolutifs importants comme l’existence de « grades » et de lignes latérales hautement spécialisées, les phénomènes d’évolution en mosaïque et tous les facteurs de causalité en évolution. À quelle vitesse diverge une nouvelle branche, dans quelle mesure elle change relativement à son « groupe frère », combien de caractères supplémentaires elle acquiert, quelle nouvelle zone adaptative a-t-elle envahi, etc., toutes ces questions sont rarement posées pas les cladistes. En ne considérant que la distance généalogique le cladiste se comporte comme s’il admettait que toutes les lignes divergent de façon équivalente et que la distance généalogique correspond à la distance génétique. En prétendant que le branchement est le seul processus historique conséquent, il refuse que les autres aspects du changement évolutif puissent être reconnus en tant que « processus historiques », comme la vitesse d’évolution, la radiation adaptative, l’occupation de nouvelles zones adaptatives, l’évolution en mosaïque, et de nombreux autres phénomènes macroévolutifs.
64 Les deux composants de la phylogénie sont potentiellement de même importance pour le taxinomiste évolutionniste, et les deux doivent être considérés judicieusement lors de la construction de la classification. Le clivage, tout comme le changement phylétique, ont lieu simultanément au cours de l’évolution mais dans la plupart des groupes l’un ou l’autre des deux processus domine pendant une période géologique donnée. Quand il y a un clivage massif, par exemple la spéciation des 50 000 ou 100 000 espèces de charançons (Curculionidae), ou des milliers d’espèces de Drosophila, la divergence phylétique est relativement insignifiante. Parmi les invertébrés et plus spécialement parmi les arthropodes, il existe de nombreux taxons où la spéciation abondante s’est produite sans aucune dérive dans une nouvelle zone adaptative. Toutes les espèces de ces groupements sont des variations répétées autour d’un seul thème. Le contraste est fort avec les épisodes de l’histoire du monde aussi mémorables que l’origine des plantes vasculaires, des angiospermes, des cordés, des vertébrés, des tétrapodes terrestres, des reptiles, ou des oiseaux.
65 Rensch (1947), Huxley (1942, 1958) et Simpson (1959b, 1961), en particulier, ont souligné l’importance de ces niveaux d’adaptation, désignés par Huxley comme des grades. Tous les membres d’un grade sont caractérisés par un complexe adaptatif bien intégré. Le succès évolutif d’une lignée phylétique vers une nouvelle zone adaptative (et à l’intérieur de cette zone) est caractérisé par l’acquisition pas à pas (évolution en mosaïque) d’une série de nouveautés pour qu’elle s’adapte (ainsi que ses descendants) à sa nouvelle position dans l’écosystème. Ultérieurement, le nouveau type basique de cette ligne phylétique peut n’éprouver que peu de changement évolutif mais expérimenter au contraire une importante radiation adaptative qui résulte en une spéciation intense et des modifications variées à l’intérieur du thème adaptatif de base du grade. Nous connaissons dans l’histoire des vertébrés de nombreux cas de formation de nouveaux grades ayant connu le succès, comme les requins, les poissons osseux, les amphibiens, les reptiles, les oiseaux et les mammifères. Chacun d’entre eux est caractérisé par un certain type d’adaptation à l’environnement (Bock 1965), quel que soit le nombre de subdivisions cladistiques dans le grade. Une grande perte d’information résulte de l’ignorance de la composante adaptative de l’évolution exprimée par le concept de grade et de l’attention aux seuls clivages des lignes. Mais c’est précisément ce que font les cladistes.
66 En réalité l’existence de grades modestes et de grades importants est l’un des phénomènes phylogénétiques les plus intéressants, même s’il s’agit d’un phénomène que nous n’arrivons toujours pas à comprendre parfaitement. Pourquoi existe-t-il si souvent une telle uniformité de type dans un taxon supérieur ? La diversité en espèces des perroquets est très riche mais tous, du petit lori de Wilhelmina ou lorikeet au cacatoès ou au grand ara, restent des perroquets caractéristiques. C’est le cas de la plupart des taxons supérieurs, sinon tous. Les reptiles représentent un grade bien défini entre le niveau amphibien et celui des deux dérivés des reptiles, les oiseaux et les mammifères.
67 Dans ses différentes publications, Simpson (1953, 1959b, 1961) a discuté à chaque fois de la différence entre les grades et les clades. Les crocodiles ont un ancêtre commun avec les oiseaux plus récents que celui qu’ils partagent avec les lézards. Ils appartiennent par conséquent au même clade que les oiseaux mais n’appartiennent pas au grade avien, ils appartiennent plutôt à celui des reptiles. Auquel de ces deux aspects de l’évolution doit-on donner la primauté ? Il y a littéralement des milliers de dilemmes semblables dans l’évolution des animaux et des plantes. Par exemple, les grands singes africains (Pan) ont un ancêtre en commun avec l’homme (Homo) plus récent que celui qu’ils partagent avec l’orang (Pongo). Néanmoins Pan appartient au même grade que Pongo, très différent de celui représenté par l’homme. Mieux sont connues les archives fossiles, plus on rencontre de tels dilemmes. Pour le taxinomiste évolutionniste l’existence de grades semble souvent plus significative et biologiquement plus sensée que le seul clivage des lignes phylétiques. Le point jusqu’où les cladistes déprécient la signification biologique des grades est illustré par un commentaire de Brundin (1972 : 111) qui désigne les groupes comme celui des reptiles d’« abstractions non temporelles ».
68 Il y a deux raisons, pour un cladiste, au fait de négliger délibérément la divergence évolutive. La première est que ce facteur ne peut pas être mesuré avec précision de façon non équivoque. De fait, on ne peut habituellement inférer les vitesses et le degré de divergence évolutive que par extrapolation et autres méthodes indirectes. Mais une pondération appropriée (Mayr 1969 : 220-228) permet des inférences probabilistes qui ont du sens, qui, quoique pas très précises, ont plus de valeur que le conseil d’ignorer tout simplement la divergence évolutive. La seconde est que les cladistes semblent penser qu’il faut faire un choix pour délimiter les taxons : ou bien les fonder sur les points de branchement ou bien sur le degré de divergence évolutive. Ils n’arrivent pas à saisir qu’utiliser les deux sources d’information augmente d’autant l’information.
69 Quand ils se voient critiqués pour leur négligence de la divergence évolutive, les cladistes essaient de se défendre en citant la règle de déviation de Hennig. « Quand une espèce se subdivise, l’une des deux espèces filles tend à dévier de l’espèce souche commune plus fortement que l’autre » (Hennig 1966 : 207). Cette règle, disent les cladistes, prouve qu’ils n’ignorent pas l’évolution phylétique. Plusieurs aspects de cette règle sont remarquables. Le premier est en totale contradiction avec l’affirmation de Hennig selon laquelle le clivage est le seul processus historique de la phylogénie. Une déviation non symétrique est un processus historique qui – en tant que tel – est indépendant du clivage. Le second est que la règle ne semble pas jouer le moindre rôle dans la construction des classifications des cladistes, même si Hennig a énoncé la règle de déviation dès 1950 (Hennig 1950 : 111) et l’a confirmé en 1966 et en 1969 (Hennig 1969 : 43) et même si elle a été en principe adoptée par la plupart des cladistes (par exemple Brundin 1972 : 108). Le schéma qui illustre la règle de déviation (Hennig 1950, Fig. 25) est l’un des rares schémas de Hennig où les angles des différents clades ne sont pas égaux. On a presque l’impression que le but même de la règle de déviation est de permettre aux cladistes de se défendre de l’accusation d’ignorance de la divergence évolutive. Les conséquences de la règle de déviation sont totalement négligées. Elles seraient d’autant plus apparentes si le processus de déviation non égale apparaissait à chaque pas de la spéciation. Il deviendrait alors évident à quel point la divergence phylétique est importante. Par exemple, si l’un des nouveaux « groupes frères » ressemble beaucoup au groupe parent, ou même est identique (comme l’indique Hennig 1966 : 59), alors que l’autre dévie fortement, la terminologie « groupes frères » ne peut plus être appliquée puisque l’on a dans ce cas un groupe parental qui se prolonge et d’où est issu un groupe fils (Hennig 1966, Fig. 14, 15). Mais au-delà de cette objection formelle il en est une de bien plus sérieuse. Si une forte accélération de vitesse de divergence apparaît dans une branche (un groupe frère) tandis que les branches subséquentes de l’autre groupe frère ne divergent que très peu, il en résulterait avec le temps un système non balancé au point d’annihiler complètement l’utilité du cladogramme dichotomique. De tels branchements asymétriques apparaissent très fréquemment au cours de l’évolution et peuvent être facilement intégrés dans la classification des taxinomistes évolutionnistes mais ne peuvent l’être que bizarrement (si même ils le sont) dans la classification cladistique.
70 Quelques disciples de Hennig ont repéré ces contradictions. Schlee (1971 : 5, 30, 37) voit clairement que l’on peut se passer de la règle de déviation : « Elle n’offre pas un argument en faveur de la méthode de Hennig ni un prérequis pour travailler selon les principes de Hennig. » Schlee (1971 : 6) ajoute une interprétation mystérieuse de Hennig lui-même : « La “règle de déviation” doit être spécialement comprise dans un sens généalogique mais non dans un sens morphologique-biologique. » En réalité, c’est l’inverse qui est vrai : dans un sens strictement généalogique il ne peut y avoir de déviation. S’il y a une déviation inégale cela doit être dans « un sens morphologique-biologique ». Une telle déviation morphologique est la situation normale en phylogénie. Quand il y a subdivision de lignes phylétiques, presque invariablement une ligne divergera plus rapidement et plus massivement que l’autre ; en fait, l’une d’elles peut ne pas changer du tout. L’absolue négligence de l’apparition fréquente de ce processus est l’une des erreurs fatales de la transcription de l’analyse cladistique dans une classification.
71 Comme la reconnaissance des grades dans la classification est parfois qualifiée dans la littérature cladistique d’approche typologique, je voudrais attirer l’attention sur un usage quelque peu différent du terme typologique (typological, typologisch) dans la littérature américaine et dans la littérature allemande. Typologique au sens américain est synonyme d’essentialiste, faisant référence à l’abstraction sous-jacente eidos (essence) et la négation de l’existence de la variation. Dans la littérature phylogénétique allemande, le type (Typus), ou plan d’organisation (Bauplan), est aussi une abstraction, qui représente aussi bien le « type » ancestral que le plan d’organisation « idéal » d’un taxon majeur. Reconnaître les reptiles comme un taxon légitime signifie que l’on reconnaît un plan d’organisation reptilien généralisé, ceci est alors tenu pour une approche typologique. L’usage allemand souligne la philosophie typologique de la morphologie idéale. L’usage allemand de typologique signifie souvent aussi « phénétique » (au sens large). La confusion entre les deux concepts de « typologique » est bien illustrée par la discussion qu’en donne Schindewolf (1967).
c. Une définition de l’espèce purement formelle
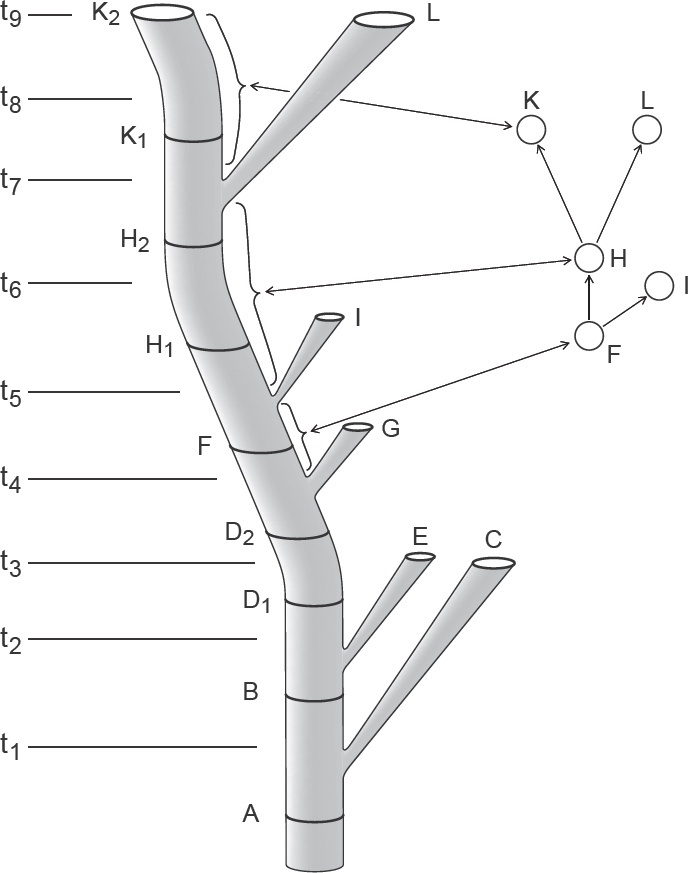
72 La définition de l’espèce par Hennig (1950) dans son traité original n’intéresse que la délimitation de l’espèce dans la dimension du temps. Une espèce est simplement la distance entre deux points de branchement dans l’arbre phylogénétique (Hennig 1950 : 111). Le concept est maintenu dans son livre de 1966 (Hennig 1966 : 56-65). Par exemple : « Les limites de l’espèce dans une section longitudinale de temps seraient en conséquence déterminées par deux processus de spéciation : le premier par lequel elle (l’espèce) émerge en tant que communauté reproductive isolée, le second par lequel les descendants de la population originale cessent d’exister en tant que communauté de reproduction homogène » (Hennig 1966 : 58). Bien que les derniers mots de cette définition aient un air de définition biologique de l’espèce, les diagrammes qui l’illustrent (en particulier sa Fig. 15, p. 60 ; Fig. 3) montrent que le concept d’espèce de Hennig est purement formel. Par exemple les espèces B et D1 ne diffèrent pas de l’espèce souche A, mais doivent être comprises comme des espèces différentes à cause des branchements respectifs de C et de E. Au contraire, l’espèce D2 est différente de l’espèce D1 (« morphologiquement » dit Hennig, mais son argument serait le même si elles étaient « reproductivement » isolées) mais doivent être nommées comme étant la même espèce car aucune branche n’est issue de cette souche.
La relation, selon Hennig, entre branchement et spéciation (d’après Hennig 1966, Fig. 15).
La relation, selon Hennig, entre branchement et spéciation (d’après Hennig 1966, Fig. 15).
73 J’attire l’attention sur le concept d’espèce de Hennig, non qu’il soit le point décisif de la théorie de Hennig (il ne l’est pas, quoi qu’en dise Hennig), mais surtout parce qu’il donne un autre exemple de la nature arbitraire et purement formelle des composants principaux de la théorie cladistique de Hennig. Le concept d’espèce de Hennig est à l’évidence si peu biologique et non réaliste qu’il a été rejeté par de nombreux auteurs critiques récents. Peters (1970 : 28-30), par exemple, réfute les formalisations de Hennig de façon efficace, en montrant que parmi toutes les propriétés possibles des espèces, leur durée dans la dimension du temps est la moins significative. Cain (1967 : 412) exprime également que le formalisme de Hennig est sans signification biologique.
74 Le concept de spéciation de Hennig est fortement dépendant de son concept d’espèce. Les différentes implications que l’on peut tirer des discussions de Hennig peuvent être formulées selon trois possibilités :
- L’espèce parente est éliminée par
- la subdivision de l’espèce mère en deux espèces filles (dichotomie), ou
- la subdivision simultanée de l’espèce mère en plus de deux espèces filles, ou
- L’espèce parente persiste
- après l’origine d’une ou plusieurs branches latérales à partir de la ligne phylétique principale essentiellement inchangée.
76 Dans son livre de 1950, Hennig n’admet que le premier point 1 et parmi les cladistes c’est toujours le cas privilégié car il s’accorde le mieux au schéma cladistique. Les cladistes n’ignorent pas le point 3 mais ont une solution formelle pour ce phénomène inadéquat. Si une espèce a donne une espèce c, l’espèce a sera appelée espèce b à partir du point de branchement afin de satisfaire le postulat cladistique, même si b n’est biologiquement pas distinguable de a. Dans les études de parenté, écrit Hennig (in Schlee 1971 : 28) : « La question de l’identité biologique de différentes espèces de différents horizons chronologiques est sans fondement. » Ceci montre à quelles extrémités mène le formalisme dogmatique de Hennig. Parce que, quoi qu’il en dise, si a et b sont biologiquement identiques, alors ce ne sont tout simplement pas des espèces différentes mais la même espèce à partir de laquelle l’espèce c s’est branchée à un moment donné. Et ceci est d’une importance capitale pour la discrimination entre les espèces sœurs et filles. La solution de Hennig est terriblement trompeuse. La production d’une branche latérale, une nouvelle lignée phylétique, ne change pas l’espèce parente.
Dichotomie ou pas
77 Les études modernes de la spéciation nous permettent de savoir à quelle fréquence les situations 1 et 2 apparaissent. Tout nous indique que le cas de la simple dichotomie en deux espèces filles n’est pas la règle. Les espèces polytypiques ont, le plus souvent, plus de deux sous-espèces. Un meilleur exemple est donné par les super-espèces qui consistent en groupes d’allo-espèces. En Amérique du Nord, au moins 48 des 126 super-espèces d’oiseaux (= 40%) ont plus de deux allo-espèces (Mayr & Short 1970). En Mélanésie du Nord, parmi 94 super-espèces 61 (= 65%) ne sont pas dichotomiques, contenant de trois à 13 allo-espèces (Mayr & Diamond sous presse). Le « standard » dichotomique est également réfuté par tous les genres riches en espèces et par la fréquence des groupements d’espèces jumelles. Ce que l’on découvre le plus souvent dans les études de spéciation est que l’espèce mère ne subit que relativement peu de changement évolutif tandis que de nombreuses espèces filles en dérivent à la périphérie. Hennig lui-même est au courant de cette situation puisqu’il montre la variation géographique du lézard Dendrophis pictus où la vaste population centrale est entourée de six populations périphériques isolées (Hennig 1966 : 61, Fig. 16). Les biologistes ont de plus en plus constaté que le bourgeonnement périphérique est le processus de spéciation le plus fréquent même si la plupart de ces espèces filles ont une durée de vie très courte. Dans ces conditions j’ai du mal à comprendre la logique de l’assertion de Nelson (1971 : 374) : « L’utilisation de la spéciation dichotomique en tant que principe méthodologique est requise avant qu’une hypothèse de spéciation multiple ne soit, même provisoirement, acceptée. »
78 La différence entre le clivage et le bourgeonnement peut n’apparaître que comme tenant purement de la sémantique, s’il ne s’agissait pas du fait que les cladistes tirent constamment leurs conclusions de la cohérence de la dichotomie. Darlington (1970 : 2-4) a publié une critique incisive. Quelques cladistes commencent même à abandonner le principe de la dichotomie obligatoire (par exemple Schlee 1971 : 27) mais ils n’en ont pas tiré toutes les conséquences vis-à-vis de leur théorie des groupes frères. Si un clivage non dichotomique produit trois ou quatre lignes phylétiques indépendantes, cela signifie que chacun des groupes qui en résultent a deux ou trois différents groupes frères plutôt qu’un seul. Dans de telles circonstances, de nombreuses discussions dans la littérature cladistique à propos des groupes frères sont sans objet.
79 Dans Stammesgeschichte der Insekten, Hennig (1969) reconnaît de nombreux taxons supérieurs qui contiennent plus de deux groupes frères. Il souligne cependant que c’est un arrangement provisoire, acceptable « aussi longtemps que les relations de parenté exactes sont incertaines ».
80 La place de l’extinction est grandement sous estimée dans les constructions cladistiques. De fait, les paires de taxons apparentés (les groupes frères de Hennig) sont fréquentes. Leur existence est habituellement liée à l’extinction de nombreuses lignes phylétiques plutôt qu’à un processus particulier de spéciation, c’est-à-dire le clivage d’une espèce mère en deux espèces filles. Il n’y a guère de preuves que ce processus ait souvent lieu, et aucune ne soutient qu’il soit le processus universel de la phylogénie.
d. Le mode d’origine des taxons supérieurs
81 Le cladiste est forcé de proposer un mode d’origine des taxons supérieurs irréaliste à cause de sa théorie phylogénétique. Puisqu’il pense que le branchement est le seul processus phylogénétique, il doit reconnaître deux propriétés à ses points de branchements : ils sont l’origine de nouvelles espèces et aussi de nouveaux taxons supérieurs. Cette idée arbitraire ne correspond nullement aux faits, ainsi que pointé par Darlington (1970 : 2). La spéciation, c’est-à-dire l’acquisition de mécanismes d’isolement dans la reproduction entre populations et l’acquisition de nouveaux caractères apomorphes significatifs sont deux processus largement indépendants. L’étude des groupes d’espèces jumelles et la plupart des genres riches en espèces montrent que l’acquisition de l’isolement reproductif est souvent (sinon toujours) sans effet sur les critères morphologiques que le taxinomiste ou le biologiste évolutionnistes associeraient à l’origine de nouveaux taxons supérieurs. Que l’une des espèces filles acquiert au cours de la spéciation un caractère qui soit potentiellement significatif pour caractériser un nouveau taxon supérieur, est l’exception plutôt que la règle. L’apparition de nouveaux caractères apomorphes est corrélée à l’invasion de nouvelles niches et de nouvelles zones adaptatives bien plus qu’à la spéciation (Simpson 1956 b ; Bock 1965). L’existence de la dichotomie phylogénétique devient donc plus plausible si elle est déconnectée de la spéciation, car la probabilité que plusieurs lignes filles dévient simultanément vers la même nouvelle zone adaptative est faible. Elle n’est cependant pas nulle car un changement de climat et de végétation ou l’arrivée d’un nouveau prédateur peut causer la même dérive simultanée chez plusieurs lignes apparentées.
82 L’acquisition des mêmes caractères apomorphes dans différentes lignes dans un ensemble de groupes frères, disons les descendants d’une super-espèce hautement polytypique, est certainement plus fréquente et potentiellement plus problématique. L’extinction fournit le plus souvent une solution pratique à ce dilemme. L’évolution d’un nouveau taxon supérieur à partir de la ligne initiée par la nouvelle espèce est un événement ultérieur, qui correspond à un second processus phylogénétique, la divergence évolutive, et n’a rien à voir avec le clivage. C’est une formulation trompeuse que de dire que les taxons supérieurs clivent. La preuve de cette assertion est la suivante : un taxon supérieur est un assemblage collectif qui comprend de nombreuses espèces et de lignes évolutives moins nombreuses. Peu d’entre elles divergeront au point de former un taxon supérieur distinct. Mais, de temps en temps, lorsque l’une de ces moindres branches diverge tellement, elle doit être séparée en tant que taxon supérieur distinct.
83 Deux points sont importants : l’« Adam » de la nouvelle ligne phylétique appartient presque invariablement au taxon ancestral. Le premier membre de la ligne phylétique qui va mener aux oiseaux (bien avant Archaeopteryx) était probablement un dinosaure par ailleurs conventionnel, mais avec des plumes ou des écailles à allure de plumes. Est-ce qu’un seul caractère dérivé est suffisant pour lancer cet « Adam » vers un nouveau taxon qui émergera finalement de cette lignée ? Les paléontologues se sont intéressés depuis longtemps à ce problème qui a été discuté de façon très détaillée par Simpson (1961 et autres publications).
84 Ashlock (1971 : 67) a fait une suggestion subtile selon laquelle la délimitation d’un taxon ancestral vis-à-vis du taxon dérivé pourrait être établie comme suit : « Les innovations trouvées chez les membres vivants pourraient être retracées à partir des fossiles et la coupure être placée quelque part entre la première apparition de l’un de ces caractères et la première apparition de l’ensemble de ces caractères […]. L’assignation spécifique de la limite pourrait être déterminée phénétiquement, en pondérant les caractères si besoin, comme pour établir que le groupe dérivé a atteint le grade. » Il admet que le manque de préservation des parties molles et une absence d’archives fossiles rendent ce but difficile à atteindre, sinon impossible, mais sur le principe, c’est certainement la procédure appropriée.
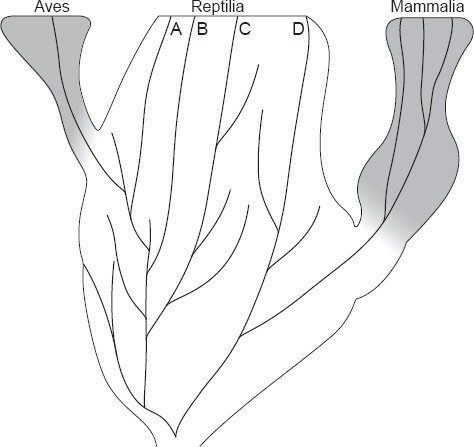
85 L’autre conclusion importante est que l’origine d’une branche latérale n’a pas de conséquence évolutive sur la branche principale (sauf en cas de compétition). Par exemple, la classe des Reptilia, un grade de tétrapodes bien caractérisé, existe depuis le Carbonifère et dont quatre ordres survivent. À un moment dans le Trias, une des nombreuses branches latérales reptiliennes (les cynodontes parmi les thérapsides) a évolué en mammifères et un peu plus tard une autre branche a donné naissance aux oiseaux (Fig. 4). Une application rigide de leur dogme force les cladistes à briser le grade reptilien en nombreuses « classes » séparées et à désigner certaines lignées reptiliennes comme le « groupe frère » des oiseaux et des mammifères. Le fait que personne ne placerait les crocodiles en dehors des reptiles si les oiseaux n’existaient pas, montre combien ce procédé est artificiel et arbitraire. L’unité essentielle des reptiles est illustrée au mieux par l’argumentation continuelle des paléontologues selon laquelle les ordres des reptiles sont mutuellement les plus proches parents.
L’indépendance de l’émergence des grades avien et mammalien vis-à-vis du pattern de branchement du grade reptilien. A (= les crocodiliens) appartient cladistiquement aux Aves, mais il est toujours un membre caractéristique du grade reptilien. L’origine des oiseaux et des mammifères n’affecte pas la catégorisation des branches reptiliennes dont elles sont issues.
L’indépendance de l’émergence des grades avien et mammalien vis-à-vis du pattern de branchement du grade reptilien. A (= les crocodiliens) appartient cladistiquement aux Aves, mais il est toujours un membre caractéristique du grade reptilien. L’origine des oiseaux et des mammifères n’affecte pas la catégorisation des branches reptiliennes dont elles sont issues.
86 L’une des difficultés majeures de la cladistique vient de l’hypothèse selon laquelle tout nouveau taxon supérieur implique la disparition du taxon parent. Même s’il existe une quantité de preuves que de nombreux taxons supérieurs, sinon tous, sont des dérivés latéraux (branches latérales) à partir d’autres taxons qui ont existé bien avant et ont persisté bien après le clivage, le cladiste, à cause de son singulier formalisme, est forcé de prétendre de façon irréaliste : « L’espèce souche n’appartient pas à l’un ou l’autre groupe (fils) (auquel elle a donné naissance) : elle ne peut pas être assignée à aucun des deux » (Hennig 1969 : 33).
87 On peut illustrer à quel point l’approche cladistique est insoutenable par l’étude d’un groupe très diversifié, qui existe toujours et est source de spéciations. La famille actuelle des Drosophilidae offre un tel exemple (Fig. 5). Dans ce taxon supérieur, nous possédons une information tellement riche à partir de la morphologie, des chromosomes, du comportement et d’autres caractéristiques, que l’on peut construire la phylogénie probable avec un degré de fiabilité très élevé. Le dendrogramme montre que plusieurs genres spécialisés prennent leur origine de divers endroits du dendrogramme de Drosophila. L’origine d’une ligne latérale spécialisée, pour laquelle on reconnaît le statut de genre, n’affecte en rien le statut taxonomique de la branche principale de Drosophila qui se poursuit en tant que genre Drosophila. Ce n’est pas un cas unique. L’origine de multiples branches latérales, chacune se spécialisant dans différentes directions, tandis que le groupe souche majeur persiste essentiellement inchangé, est une situation très courante en phylogénie.
L’origine des genres spécialisés à l’intérieur de l’arbre phylétique de Drosophila (d’après Throckmorton 1965).

L’origine des genres spécialisés à l’intérieur de l’arbre phylétique de Drosophila (d’après Throckmorton 1965).
2. Une conception trompeuse de la catégorisation
88 Le processus de classification consiste essentiellement en deux étapes, 1. le groupement des taxons inférieurs (habituellement les espèces) en taxons supérieurs, et 2. l’assignation de ces taxons à des catégories appropriées dans la hiérarchie taxinomique (catégorisation). Les considérations qui commandent à ces deux processus sont fort différentes l’une de l’autre (Mayr 1969, chap. 10). Les cladistes, en fonction de leur idée de base selon laquelle la phylogénie est une procédure unique (qui consiste seulement dans le branchement), affirment que la classification est une procédure à une seule dimension et que le groupement des taxons donne simultanément leur rang. Traditionnellement, le rang des taxons est déterminé par divers critères comme le degré de différence, l’originalité de la zone adaptative occupée, ou l’importance de la radiation adaptative qui s’y déploie (Mayr 1969 : 233). Selon la cladistique, le rang est donné automatiquement par la date d’origine, et le même rang doit être donné aux groupes frères.
89 Cette conclusion erronée est renforcée par la fréquente confusion qu’entretient Hennig entre taxon et catégorie. En 1950 une telle confusion était excusable car jusqu’à cette époque on n’avait pas proposé de distinction terminologique. En 1966 la confusion n’était plus défendable en raison de la différence largement expliquée par de nombreux auteurs. Cependant même en 1966, Hennig (1966 : 77-83) parle de la « réalité et l’individualité » des catégories, quand il s’agit de taxons, et cette confusion obscurcit terriblement les procédures de catégorisation. Tout le processus de classification est fondé sur la distinction claire du taxon et de la catégorie : dans la nature, les groupes zoologiques (taxons) ont une « réalité » parce que les noms des taxons comme oiseaux, papillons, ou chauves-souris, sont des noms non ambigus pour des groupes facilement distinguables. Il est tout aussi évident que la catégorisation de ces taxons dans la hiérarchie linnéenne des catégories taxinomiques est plutôt arbitraire, et souvent très controversée. Ce qu’un auteur va considérer comme une tribu, un deuxième va l’appeler sous-famille et un troisième une famille, voire une superfamille (voir ci-dessous).
90 Malheureusement Hennig croit (ou a cru au début) que la connaissance des points de branchement pourrait lui permettre de déterminer le rang d’un taxon. Il utilise deux critères : l’âge géologique du point de branchement (qui mène au groupe frère le plus proche) et le nombre de points de branchement ultérieurs. Il l’a déclaré à de nombreuses reprises dans ses écrits, sans réserve. Par exemple, dans la hiérarchie les taxons « sont subordonnés les uns dans les autres en fonction de la distance temporelle entre les origines et le présent ; la séquence de la subordination correspond au “degré d’ascendance commune” » (Hennig 1950 : 83). Et « dans le système phylogénétique […] l’ordre absolu des rangs ne peut être indépendant de l’âge du groupe puisque […] la coordination et la subordination des groupes sont mises en place par la définition de l’âge relatif d’origine » (Hennig 1966 : 160).
91 Les deux critères de Hennig ont été souvent critiqués. Peters (1970), par exemple, reconnaît que parmi deux lignes évolutives qui évoluent à la même vitesse, l’une peut donner un grand nombre de branches latérales tandis que l’autre peut rester d’une singularité monolithique. Attribuer à la ligne qui produit des branchements fréquents un plus haut rang qu’à la ligne qui n’en produit pas, n’aurait aucun sens. En fin de compte, le branchement et la divergence évolutive sont deux processus indépendants. Hennig et quelques-uns de ses disciples parlent à l’occasion maintenant « d’âge chronologique relatif » à la place de « l’âge géologique absolu » mais en 1966 Hennig maintenait que l’emplacement des points de branchement sur l’échelle géologique indiquait le rang de la catégorie (Fig. 6).
L’assignation de la catégorie sur la base de l’âge géologique absolu de l’espèce souche (d’après Hennig 1966 : 187 ; Fig. 54).

L’assignation de la catégorie sur la base de l’âge géologique absolu de l’espèce souche (d’après Hennig 1966 : 187 ; Fig. 54).
92 En conséquence, les taxons qui ont leur origine dans le Précambrien sont catégorisés comme des phylums ; entre le Cambrien et le Dévonien comme des classes ; entre le Mississippien et le Permien comme des ordres ; entre le Trias et le Crétacé inférieur comme des familles ; entre le Crétacé supérieur et l’Oligocène comme des tribus ; et au Miocène comme des genres. Hennig (1966 : 187) continue : « Ainsi les mammifères devraient être appelés un ordre […]. [L]es Marsupialia et les Placentalia devraient être descendus au rang de la famille, et les “ordres” des Placentalia devraient être des tribus. » On rencontre des affirmations semblables chez d’autres cladistes. Par exemple : « Les espèces doivent être catégorisées 1) en raison de leur âge relatif d’origine ou 2) de telle façon que les groupes frères aient le même rang » (Nelson 1972 : 366). « Age relatif » est une expression fréquente dans la littérature cladistique mais personne n’a jamais proposé une méthode opérationnelle pour déterminer l’âge relatif.
93 Si l’on voulait vraiment catégoriser les taxons sur la base de leur date d’origine (géologique), on serait obligé d’adopter un système exclusivement non balancé. Puisque l’origine du genre Lingula est plus ancienne que celui de la classe Aves ou de la classe Mammalia on devrait placer Lingula (et d’autres genres contemporains qui survivent aujourd’hui) dans une catégorie plus élevée que celle des oiseaux et des mammifères. Même un cladiste apparaîtrait inconséquent d’opter pour de telles extrémités. Les discussions entre Griffiths (1972 : 10) et Crowson (1970 : 250-254) montrent ces difficultés.
94 Curieusement Hennig a mentionné la classification des parasites à l’appui de la méthode cladistique (Hennig 1950 : 261-269). Il confirme que « l’on doit donner le même rang systématique au groupe hôte et au groupe de parasites […] si le groupe de parasites, qui est propre à un certain groupe hôte, a la même origine temporelle, peut-être en lien avec le fait que l’espèce souche du groupe de parasites vivait en parasite du groupe hôte » (Hennig 1950 : 265). Sauf que, comme Osche (1961) l’a avancé, la chronologie des parasites et de leurs hôtes réfute en fait la méthode cladistique. Il est hautement probable que l’origine de nombreux parasites de vertébrés se situe à la même période géologique que leurs taxons hôtes. Cela voudrait dire, si l’on suit strictement Hennig, qu’un genre de cestode devrait être élevé au rang familial ou ordinal s’il parasite une famille ou un ordre de vertébrés, ou, réciproquement, que le taxon hôte soit déplacé au rang générique. Cela voudrait dire aussi que chez les nématodes, la superfamille des Ascaroidea devrait être située au même rang que la classe des Cestoda parce que l’on pense que l’espèce souche des deux taxons a envahi les vertébrés au même moment.
95 Plus les phylogénies sont étudiées soigneusement plus on recense des difficultés pour appliquer les principes hennigiens de catégorisation. Les différents moyens auxiliaires évoqués par Hennig n’arrangent pas la situation. En assumant que le branchement est un processus régulier, avec une même vitesse approximative, Hennig pense que le nombre d’espèces contenues dans un taxon est une mesure approximative de son âge : « Les groupes monophylétiques pourvus d’un grand nombre d’espèces ne peuvent pas être très jeunes » (Hennig 1966 : 182). Des recherches évolutionnistes récentes ont clairement montré que cette conclusion ne tient pas. La corrélation entre l’âge d’un taxon et le nombre d’espèces qu’il contient est extrêmement faible. Les drosophilidés d’Hawaï sont clairement un groupe monophylétique, non seulement très récent (probablement moins de 4 millions d’années) mais aussi très riche en espèces (au moins 600 à 800). La plupart des nuées d’espèces de poissons cichlidés d’Afrique sont également très récentes, remontant aux tout derniers millions d’années. La plupart des familles de rongeurs, riches en espèces, sont géologiquement plus récentes que la plupart des familles de mammifères qui possèdent deux ou trois genres. Les monotrèmes avec quatre ou cinq espèces sont aussi vieux ou plus vieux que les euthériens qui comptent plus de 3 000 espèces (Peters 1970 : 31). Des invertébrés marins bien connus remontent à 300-400 millions d’années sans avoir jamais produits d’importants groupes d’espèces.
96 Hennig et ses supporteurs prétendent que le grand mérite de la méthode cladistique est de fournir des définitions non arbitraires des catégories supérieures. Griffiths (1972 : 16), par exemple, félicite Hennig d’avoir proposé une définition logique non critiquable « en termes d’âge d’origine des espèces souches ». J’ai montré plus haut à quel point cette proposition est irréaliste et à quel point elle échoue à délivrer une procédure sensée de catégorisation. Les paléontologues des invertébrés qui peuvent illustrer les destins évolutifs extrêmement différents de différentes lignes phylétiques (dérivées d’une même espèce souche) ont fréquemment souligné que la méthode cladistique de catégorisation aboutit à des classifications déséquilibrées. De la même façon, Simpson (1961 : 142-144) a décrit à quoi aboutirait la classification des vertébrés en appliquant ces principes. Crowson (1970 : 260) admet que « si la classification phylogénétique part habituellement des divisions dichotomiques, très inégales de surcroît, il serait nécessaire de disposer d’un bien plus grand nombre de catégories [et de noms de taxons] que pour les anciens systèmes “formels” ». Simpson a entièrement raison lorsqu’il dit qu’une telle classification est « totalement impraticable ». Hennig lui-même a désormais réalisé (Hennig 1969 : 10) que cette méthode entraîne une échelle de catégorisation absurde et a abandonné dans son livre sur la phylogénie des insectes toute tentative pour fournir des rangs aux taxons supérieurs. Mais de quelle méthode s’agit-il si elle ne peut pas catégoriser les taxons supérieurs ?
3. Une négligence opérationnelle de faits évidents
97 De nombreux aspects de la phylogénie posent des problèmes à la cladistique. Hennig et ses partisans ne sont pas ignorants de ces faits et les discutent librement à l’occasion. Cependant, lorsqu’ils établissent leur classification, ils ignorent ces difficultés, ou, tout au moins, ils n’offrent pas d’instruction opérationnelle pour les surmonter. J’y ai déjà fait allusion à propos de la discussion sur la « règle de déviation » (voir ci-dessus) où la divergence inégale des lignes phylétiques est reconnue sans en tirer les conséquences pour la classification. Les autres faits négligés sont les suivants.
a. La difficulté de déterminer la direction des séquences évolutives
98 Les taxons sont construits par Hennig sur la base des synapomorphies (caractères dérivés). Pour chaque séquence de caractères il faut donc déterminer la direction du changement évolutif. Il est parfois facile de le faire ; dans de nombreux autres cas c’est très difficile. Hennig est bien conscient de ce problème et a édicté quatre règles (Hennig 1950 : 172 et suiv.) qui permettent de déterminer parmi plusieurs caractères lesquels sont ancestraux et dérivés. La plupart de ces règles, reformulées par Peters & Gutmann (1971 : 242) sont fondées sur des pratiques qui remontent à l’époque de Darwin, voire avant. Peters & Gutmann (1971 : 256) ont souligné avec raison que tout formalisme en recherche phylogénétique (cladistique incluse) tend à donner des conclusions superficielles et, souvent, peu fiables. La reconstruction phylogénétique doit être fondée sur une analyse fonctionnelle et sur sa signification adaptative, telle que pratiquée en morphologie évolutionniste (Bock 1965, 1969). Je renvoie à l’essai de Peters & Gutmann (1971) pour une perception très aigüe de cette faiblesse de la cladistique. Leurs considérations montrent bien que l’importance de l’innovation adaptative est absolument cruciale pour la classification, un fait régulièrement ignoré par les cladistes.
b. La discrimination entre parallélisme et convergence
99 La possession commune du même caractère par deux taxons différents peut être due à l’une des quatre causes (Mayr 1969 : 202).
- Ressemblance plésiomorphe. Le partage d’un caractère avec un ancêtre (voir la littérature cladistique pour plus de précisions).
- Ressemblance synapomorphe. Le partage unique de caractères dérivés à partir d’une espèce souche ; par exemple tous les oiseaux, mais aucun autre organisme, ont des plumes.
- La ressemblance due au parallélisme. Les caractéristiques produites par un génotype hérité d’un ancêtre commun.
- La ressemblance due à la convergence. Il peut s’agir aussi bien d’acquisitions convergentes (l’aile chez les ptérodactyles et les chauves-souris) ou de pertes convergentes (l’absence de pattes chez les serpents et les amphisbènes).
101 Distinguer ces quatre possibilités n’est pas si facile qu’il peut paraître à première vue, car tout ce à quoi peut accéder celui qui étudie la phylogénie est le phénotype, alors que la phylogénie consiste dans le changement des génotypes.
102 La recette traditionnelle de la discrimination entre parallélisme et convergence est l’analyse de l’homologie. Les cas où l’homologie entre les caractères peut être établie, c’est-à-dire la dérivation à partir du caractère équivalent chez l’ancêtre commun, sont le résultat de l’évolution parallèle. Les ressemblances entre les caractères non homologues sont le résultat de la convergence (Bock 1963, 1967). Suivre cette recette est cependant très difficile en pratique non seulement parce que l’établissement de l’homologie est gros de complications (Hull 1967), mais aussi parce que dans des parents éloignés la ligne de séparation entre parallélisme et convergence n’est pas nette (pour une discussion sur le parallélisme, voir également Simpson 1961 : 103-106). Osche (1965) a publié une analyse pertinente des difficultés dues aux potentialités du génotype caché.
103 Les cladistes ne font pas très attention à ces différentes possibilités : « Lorsque l’on décide si les caractères correspondants chez plusieurs espèces sont des synapomorphies, des convergences, des homologies ou des parallélismes, nous devons déterminer si le même caractère était déjà présent dans l’espèce souche exclusive des porteurs de ces caractères identiques » (Hennig 1966 : 120). Contrairement à ce que dit Hennig cela ne permet pas de distinguer les quatre possibilités mais seulement de discriminer les synapomorphies d’un côté et, de l’autre, les trois autres possibilités. Ici et en d’autres occasions (par exemple Hennig 1966 : 121) Hennig affirme carrément son désintérêt en matière de distinction entre parallélisme et convergence (voir également Peters 1972 : 168). Or cette distinction est d’une importance cruciale afin de distinguer le degré de parenté entre les taxons.
104 Les taxons sont classifiés en cladistique sur la base de la présence ou de l’absence de caractères dérivés. De fait, Hennig a répété avec insistance que rien ne compte plus en classification que de tels caractères. Malheureusement seule une partie du génotype est exprimée dans le phénotype visible et la partie cachée du génotype est souvent aussi importante pour l’évolution future d’une ligne phylétique que ce qui est révélé par le phénotype visible. Hennig (1950 : 176) a montré lui-même que le potentiel pour avoir des yeux pédonculés est largement réparti parmi les diptères acalyptrates mais n’est réalisé que chez des espèces et des genres disséminés ici et là. Une articulation mandibulaire secondaire s’est mise en place indépendamment au moins 14 fois dans la classe des Aves (Bock 1969). Dans le genre Drosophila la manifestation répétée de potentialités dissimulées est extrêmement fréquente (Throckmorton 1962, 1965). On trouve difficilement un taxon supérieur dans lequel de tels caractères dérivés n’apparaissent pas de façon disséminée dans le système. Dans tous ces cas la présence ou l’absence d’un caractère n’est en rien liée à la présence ou l’absence du caractère chez l’ancêtre commun. À vrai dire, le génotype de l’ancêtre commun a le potentiel pour développer ce caractère mais sa réalisation est imprévisible.
105 Quand des tendances émergeantes se réalisent pleinement de façon indépendante chez plusieurs caractères il n’y a pas moyen de déterminer la séquence d’apparition des caractères dérivés dans les différentes lignes, ainsi que la séquence de branchement des différentes lignes apparentées. On peut le montrer dans un schéma (Fig. 7). Le seul moyen pour un cladiste de résoudre le problème est de dire que ce dernier disparaîtra en prenant en compte un plus grand nombre de caractères. Malheureusement ce n’est pas toujours vrai. On ne dispose habituellement pas de caractères suffisamment nombreux, et quand on a peu de caractères il faut prendre des décisions à partir de pondérations soigneuses que l’on a peine à distinguer de décisions arbitraires. Dire qu’il n’y a pas vraiment de différence entre convergence et parallélisme (Hennig 1966 : 121) est signe d’un refus à prendre en compte ce qui se révèle comme une des faiblesses majeures du système cladistique. Ce ne sont pas de purs problèmes théoriques. L’effort récent d’Inger (1967) et de Kluge & Farris (1969) pour obtenir une phylogénie satisfaisante des anoures implique dans une large mesure des décisions concernant le degré de primitivité de différents caractères.
L’information contradictoire fournie par différents jeux de caractères liés à l’évolution en mosaïque. Selon le jeu de caractères 1 les lignées B et C forment le groupe frère de A ; selon le jeu de caractères 2 les lignées A et B forment le groupe frère de C.
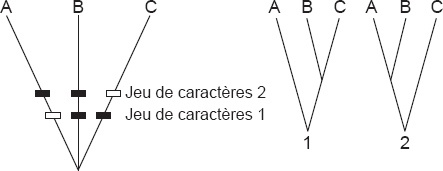
L’information contradictoire fournie par différents jeux de caractères liés à l’évolution en mosaïque. Selon le jeu de caractères 1 les lignées B et C forment le groupe frère de A ; selon le jeu de caractères 2 les lignées A et B forment le groupe frère de C.
106 Griffiths (1972 : 24-26) a consacré un long développement sur la question de savoir si la convergence pose ou non un problème à la cladistique. Il a certainement raison de penser que des caractères (ou combinaison de caractères) hautement complexes ne sont pas susceptibles d’être acquis indépendamment dans les lignes phylétiques non apparentées. (Pour une discussion du critère de complexité voir Peters 1972 : 168-170). Cependant des caractères (même complexes) peuvent être perdus indépendamment dans différentes lignes et des caractères relativement simples peuvent être acquis par convergence. Malheureusement Griffiths (1972 : 25) a mal interprété Darwin dans la phrase qui suit : « On a élevé des doutes sur la validité de la distinction faite par Darwin entre caractères “adaptatifs” et “non adaptatifs” à des fins d’évaluation évolutive, et par conséquent je n’applique pas ce critère. » En fait Darwin n’a pas fait une telle distinction. Il a simplement parlé de spécialisations ad hoc qui ont véritablement moins de poids phylétique et peuvent être acquises par convergence. C’est ce que Cracraft (1972) n’a pas bien perçu dans sa discussion récente sur l’évolution des ratites. Si plusieurs groupes d’oiseaux coureurs perdent indépendamment leur capacité à voler, acquièrent une grande taille et se spécialisent exclusivement à la course, on devrait s’attendre à ce que ces groupes aient acquis le complexe de caractères ratite même si ces caractères étaient absents chez l’espèce souche de cet ensemble. Ceci est totalement indépendant de la question de savoir si les familles d’oiseaux qui ont perdu la capacité de voler étaient formées de genres étroitement apparentés ou de genres seulement apparentés de façon lointaine. La même objection peut être potentiellement avancée vis-à-vis de l’adaptation à la plongée d’Hesperornis, des grèbes et des plongeons. Les arguments de l’école cladistique n’atténuent pas l’avertissement de Darwin sur le fait qu’il convient de ne pas trop se fonder sur des spécialisations ad hoc (voir également Mayr 1969 : 220, 223).
107 Hennig (1966 : 121) croit que l’on peut distinguer entre convergence (et parallélisme) et synapomorphie en « prenant en compte autant de caractères que possible ». C’est en principe vrai mais souvent impossible à faire. Schlee (1971 : 23) ajoute des critères supplémentaires qui sont, de fait, très utiles. Cependant, dès qu’on étudie de près les phylogénies telles qu’elles sont construites, il devient évident qu’on en tire rarement une décision non équivoque. En raison de l’aspect lacunaire des archives fossiles d’insectes comparées à celles des mammifères, les taxinomistes d’insectes ne sont pas prévenus autant qu’ils le devraient de la fréquence du parallélisme. Par exemple, « à l’intérieur des cynodontes, le groupe le plus avancé (au sein des reptiles thérapsides), de nombreux traits mammaliens (comme l’articulation cranio-mandibulaire ente dentaire et squamosal, la perte de plusieurs générations de dents de remplacement, l’occlusion complexe, et des dents jugales bi-radiculées) sont considérées comme ayant évolué indépendamment dans plusieurs lignes phylétiques » (Crompton & Jenkins 1973 : 138). La plupart des auteurs acceptent également l’idée que l’incorporation du carré et de l’articulaire dans l’oreille moyenne a dû se produire indépendamment chez les mammifères thériens et non thériens. La fréquence de l’acquisition indépendante d’adaptations identiques chez les oiseaux, comme l’articulation mandibulaire secondaire (Bock 1959) ou les nombreuses spécialisations des différentes lignées de pics (Bock & Miller 1959), ou encore les avancées phylétiques indépendantes dans le genre Drosophila (Throckmorton 1965) illustrent les difficultés (voir Bock 1967 et Peter 1972 : 168).
108 Ce qui est le plus déconcertant est le fait que lorsqu’ils construisent leurs classifications, les cladistes ne semblent pas prendre en compte les difficultés causées par le parallélisme et la convergence. Ils les ignorent simplement.
c. Le contenu informatif des caractères ancestraux
109 Les cladistes critiquent avec raison que le seul critère de délimitation des taxons soit la « simple ressemblance » mais telle est l’attitude des taxonomistes évolutionnistes depuis longtemps. Ghiselin (1969b), Simpson (1961) et moi-même (Mayr 1969) ont régulièrement pointé du doigt les dangers de fonder entièrement la classification sur la ressemblance, en particulier (comme le proposent les phénéticiens) sur la ressemblance non pondérée. Toutefois cela ne justifie pas d’aller vers l’autre extrême qui est de rejeter toute fonction à la ressemblance lorsque l’on construit une classification. Agir ainsi aboutirait à une perte considérable d’information taxinomique. En effet, quand elle est correctement évaluée la ressemblance est un indice important, mesurant la quantité partagée de génotype et offrant la base de la détermination de l’homologie. Les cladistes ignorent complètement cela et, par voie de conséquence, ne font guère attention au fait que la rétention d’un grand nombre de caractères ancestraux est tout autant un indicateur de « relations de parenté » (définies traditionnellement) que l’acquisition commune de quelques caractères « dérivés ». On a envisagé deux extrêmes vis-à-vis de l’importance relative des caractères conservateurs (ancestraux) et avancés (dérivés). Les cladistes considèrent seulement les derniers, alors qu’il a été parfois dit que c’est l’étude des caractères conservateurs qui est la plus apte à révéler la parenté. Le taxinomiste évolutionniste est convaincu (et a travaillé ainsi depuis les 100 dernières années) que l’on doit évaluer l’information des deux types de caractères dans le but de construire une classification valide. Rien n’est plus éloigné de la vérité que de dire que « les ressemblances primitives ne contiennent pas d’information phylogénétique » (Cracraft 1972 : 383).
110 J’illustrerai cela à l’aide de deux exemples : il existe trois familles principales d’oiseaux gallinacées vivants. Parmi elles les Megapodiidae possèdent le plus grand nombre de caractères primitifs tandis que les Phasianidae possèdent le plus grand nombre de caractères dérivés. Les Cracidae d’Amérique du Sud sont intermédiaires. Ils partagent quelques caractères dérivés avec les Phasianidae mais ont en commun avec les Megapodiidae un bien plus grand nombre de caractères primitifs. Traditionnellement, on a considéré que les Cracidae étaient plus proches parents des mégapodes que des Phasianidae parce qu’ils semblent partager dans une large mesure un même génotype. À l’inverse, en tant que cladiste, Cracraft (1972 : 283) souligne que les Cracidae sont plus proches parents des Phasianidae car les deux ont en commun quelques caractères dérivés. Je considère que c’est une affirmation fallacieuse. Dans leur ensemble, les caractères des Cracidae sont, à l’évidence, plus proches de ceux des Megapodiidae que des Phasianidae. Il est important de savoir que les Phasianidae se sont branchés à partir de la même branche que celle qui mène aux Cracidae, mais cela ne représente qu’une partie des données qui permettent de construire une classification.
Deux interprétations différentes de l’assignation taxinomique de mouches éteintes. Selon l’interprétation traditionnelle (contour en trait continu) les fossiles (e) appartiennent aux Sciadoceridae (A) ; selon l’interprétation cladistique (ligne de tirets) ils appartiennent aux Phoridae (E). a-h = lignes secondaires des Sciadoridae avec différentes potentialités évolutives, la plupart (c, d, f, g, h) étant éteintes.

Deux interprétations différentes de l’assignation taxinomique de mouches éteintes. Selon l’interprétation traditionnelle (contour en trait continu) les fossiles (e) appartiennent aux Sciadoceridae (A) ; selon l’interprétation cladistique (ligne de tirets) ils appartiennent aux Phoridae (E). a-h = lignes secondaires des Sciadoridae avec différentes potentialités évolutives, la plupart (c, d, f, g, h) étant éteintes.
111 Citons un autre exemple que Hennig (1971 : 12-14) a utilisé pour illustrer la supériorité de l’approche cladistique (voir Fig. 8). Au Canada, l’ambre d’âge Crétacé supérieur a livré deux diptères fossiles qui montrent des relations avec deux familles actuelles, les Phoridae, une famille cosmopolite qui contient à peu près 2 500 espèces, et les Sciadoridae avec deux espèces actuelles, les deux connues dans les continents austraux. Dans laquelle des deux familles actuelles situer ces fossiles ? Les auteurs qui ont décrit à l’origine les fossiles (McAlpine & Martin 1966) leur ont reconnu une lointaine relation avec les Phoridae mais ont placé les fossiles dans les Sciadoridae parce qu’ils ressemblent plus aux espèces de cette famille qu’à tout phoridé actuel.
112 Les deux fossiles du Canada (Sciadophora et Prioriphora) ne diffèrent de façon consistante des Sciadoceridae vivants que par un seul caractère apomorphe, l’absence d’une cellule discale. Dans le cas de deux autres caractères apomorphes (absence de la cellule anale et de l’arista dorsale), les Sciadoceridae actuels sont variables. L’un des deux fossiles canadiens (Prioriphora) a acquis quelques caractères apomorphes supplémentaires ressemblant aux phoridés (par exemple la perte d’un proscutellum, un palpe élargi à la pointe, insertion de R4+5 éloignée de l’extrémité de l’aile, m1 ne rejoignant pas m2) mais pour tous ces caractères l’état plésiomorphe (ressemblant aux Sciadoceridae) est conservé chez l’autre fossile canadien (Sciadophora). Bien que les fossiles canadiens (en tant que groupe) montrent un ou deux caractères dérivés, ils sont beaucoup plus en accord dans la majorité de leurs caractères avec les Sciadoceridae vivants qu’avec les Phoridae. En particulier ils ne possèdent pas la fusion additionnelle et la réduction de la longueur de la veine r3 de l’aile ni la perte de la deuxième cellule basale. Chez les phoridés typiques, également, les coxae et fémurs sont plus gros et l’ensemble est plus puissant.
113 Dans cet exemple, et plusieurs autres que j’ai étudiés, la nouvelle classification cladistique tout entière repose sur un ou deux caractères. Ceci n’est implicitement rien d’autre que le retour de la « classification fondée sur un seul caractère ». En dépit de leur exhortation à fonder la classification sur l’holomorphe (= la totalité des caractères), pratiquement toutes les dichotomies des classifications sont virtuellement fondées sur très peu de caractères, souvent un seul. Ceci a été fort justement critiqué par Darlington (1970 : 17).
d. Évolution en mosaïque
114 Dans la mesure où sont impliqués les états ancestraux vs états dérivés, la cladistique suppose qu’une fois que l’« évolution » a « décidé » de donner à une ligne phylétique l’état primitif d’un caractère et au « groupe frère » l’état dérivé du caractère, la distinction se perpétuera pour toujours. Il existe de nombreuses situations où c’est bien le cas. Dans d’autres, et ceci est totalement ignoré par les cladistes, le potentiel pour acquérir l’état dérivé existe chez toutes les lignes sœurs issues de l’ancêtre originel ; et, comme je viens de le souligner, ceci n’est réalisé qu’irrégulièrement dans les différentes lignes sœurs, et à des vitesses différentes, ce qui mène à l’évolution parallèle. L’annonce selon laquelle « si l’on peut identifier des caractères dérivés, alors on peut construire des lignées monophylétiques » (Cracraft 1972 : 381) n’est manifestement pas justifiée.
115 Les cladistes n’ont pas pu résoudre les difficultés liées à l’évolution en mosaïque. Cela dit, il n’est pas vrai (comme il a été dit) qu’ils croient que les dichotomies créent des groupes primitifs et évolués. Ils comprennent, comme tout bon taxinomiste, que la plupart des groupes possèdent un mélange de caractères primitifs et dérivés. Même le plus primitif des groupes de mammifères vivants, les monotrèmes, possède un certain nombre de caractères dérivés.
116 En outre, le fait que de nouveaux caractères récemment acquis sont parfois perdus au cours de l’évolution subséquente, crée une difficulté supplémentaire. Une telle double apomorphie (= état primitif secondaire) mimerait une plésiomorphie. Bien que certains cladistes reconnaissent cette possibilité, il ne me vient pas que ceci ait jamais été pris en considération lors de la construction des classifications cladistiques.
Darwin et la classification
117 Les taxinomistes évolutionnistes sont convaincus depuis longtemps qu’ils suivent strictement les principes darwiniens de la classification parce qu’ils considèrent de la même façon le branchement et le changement phylétique. Comme Simpson (1961 : 52) l’a déclaré : « La taxinomie évolutionniste s’enracine explicitement et presque exclusivement dans Darwin » (voir Simpson 1959a pour une discussion plus détaillée la théorie de la classification de Darwin). Ces dernières années les cladistes ont essayé de tirer Darwin de leur côté et Nelson (1971 : 375) est allé très loin en déclarant : « S’il existe vraiment une école cladistique, Darwin est son fondateur et son principal avocat. » Nelson (1972 : 370) a mentionné la cladistique comme « la classification de Darwin-Hennig ». Existe-t-il une seule justification de cette affirmation ?
118 L’Origine des espèces de Darwin (1859) est la première publication importante qui propose l’évolution par ascendance commune, un concept tout nouveau à l’époque (et bien différent du concept d’évolution au sens de Lamarck). Dans ce livre le langage généalogique joue naturellement un rôle important. Comme les taxinomistes évolutionnistes modernes (voir ci-dessus), Darwin se fonde sur une base strictement généalogique : « [L’]arrangement […] doit, pour être naturel, être rigoureusement généalogique. » Comme n’importe quel groupe d’animaux ne peut avoir qu’une seule généalogie, comme nous l’avons vu plus haut, le postulat de Darwin est considéré par tous les taxinomistes évolutionnistes comme un axiome. Cela dit, pour Darwin, ce n’est que le premier pas car il poursuit : « [M]ais […] la somme des différences dans les diverses branches ou groupes, alliées d’ailleurs au même degré de consanguinité avec leur ancêtre commun, peut différer beaucoup, car elle dépend des divers degrés de modification qu’ils ont subis ; or, c’est là ce qu’exprime le classement des formes en genres, en familles, en sections ou en ordres » (Darwin 1859 : 420 [6]). (Voir également Ghiselin 1969a : 84.) Il est significatif que sur ce point Darwin renvoie à sa figure du chapitre 4 (face à la page 116 de l’édition originale). Il montre que les trois espèces siluriennes congénères (A, F et I) ont évolué en 15 genres modernes. Cela représente trois « groupes frères » qui ont dérivé des trois genres siluriens. Les descendants de A et de I constituent désormais des familles distinctes ou même des ordres (Darwin 1859 : 125). « Mais on peut supposer que le genre existant F14 n’a été que peu modifié, et on pourra le grouper avec le genre primitif F dont il est issu ; c’est ainsi que quelques organismes encore vivants appartiennent à des genres siluriens » (Darwin 1859 : 421 [7]). On ne peut souhaiter d’énoncé plus explicite pour réfuter l’affirmation selon laquelle Darwin était un avocat de la classification cladistique.
119 Ghiselin & Jaffe (1973) ont montré à quel point Darwin dans sa classification des Cirripedia s’est éloigné de la classification cladistique : il place Alcippe dans les Thoracica même s’il se trouve sur la souche Abdominalia ; Pachyplasma placé sur la branche qui mène aux Balanidae est inclus dans les Chthamalinae ; Pollicipes donne naissance aux Lepadidae (où il est inclus) et par diphylie aux cirripèdes dépourvus de pied (Varrucidae et Balanidae). Il n’aurait pas conçu ces classifications s’il avait adopté la définition cladistique de la monophylie. À chaque fois Darwin construit ce que les cladistes appelleraient un groupe paraphylétique.
120 À l’opposé de ce que dit Nelson, Brundin (1972 : 107) affirme que la méthode de Hennig révèle la « faiblesse de la théorie néodarwinienne actuelle ». Aucun autre cladiste ne soutient cela. Dans de nombreux endroits il apparaît que Brundin ne comprend pas la théorie darwinienne. Croit-il à une sorte d’orthogenèse quand il dit : « Le processus évolutif est nettement plus ordonné que ne l’admettent aujourd’hui les néodarwiniens » (Brundin 1972 : 119) ?
Les objections cladistiques aux méthodes de classification évolutionniste (combinée)
121 Griffiths (1972 : 16) a écrit que le groupement « combiné » (= évolutionniste) doit être rejeté car il « soulève de sérieuses difficultés logiques ». De plus, Hennig a répété qu’une systématique « synthétique » (Hennig 1971 : 11) ou « syncrétique » (Hennig 1966 : 77) est incapable d’élaborer un système cohérent (« vide la combinaison de toute valeur scientifique ») et entraîne de grosses erreurs. Cela dit, Griffiths (1072 : 15-17) est le seul auteur qui a essayé sérieusement de faire la liste de ce qu’il considère comme les défauts de la méthode évolutionniste et de les classer. Afin d’éviter tout futur malentendu, je vais essayer de réfuter certaines de ses objections.
122 Griffiths (1972 : 16) dit que je rejette tout effort afin d’arriver à « une correspondance non ambiguë entre phylogénie et classification au-dessus du niveau de l’espèce ». Au contraire, je critique la définition étroite de la phylogénie selon Hennig, qui force les cladistes à ignorer au moins la moitié de l’information fournie par la phylogénie. C’est cela mon argument, lorsque je dis que la méthode traditionnelle de classification reflète beaucoup mieux la phylogénie que la méthode de Hennig. Griffiths dit plus loin qu’il est impossible de mesurer correctement des vitesses de changement évolutif. C’est exact ! Peut-être n’est-il pas approprié de dire qu’une classification devrait exprimer des « vitesses d’évolution ». Cependant, un classificateur ne devrait pas, ne peut pas, ignorer les résultats de vitesses d’évolution très diverses. Même s’il se révélait impossible de comparer mathématiquement la (faible) vitesse d’évolution de l’espèce souche vers les crocodiles modernes et la vitesse (élevée) entre la même espèce souche et les oiseaux modernes, n’importe quel débutant peut voir que les oiseaux diffèrent de l’ancêtre commun infiniment plus que ne le font les crocodiles. Ignorer cela, parce que la mesure n’est pas (encore) exacte, ressemble à une piètre échappatoire vis-à-vis des difficultés.
123 Une troisième objection est que les évolutionnistes n’arrivent pas à donner une définition objective des catégories au-dessus du niveau spécifique. Selon Griffiths (1972 : 7-8) leur définition ne souffre pas la comparaison avec « la proposition de Hennig de définir les catégories au-dessus du niveau spécifique en fonction de l’origine de l’espèce souche des membres des taxons ». Griffiths oublie que Hennig a échangé la signification biologique contre l’espérance d’une cohérence logique. Comme Darwin l’a montré (voir ci-dessus), les descendants modernes de deux espèces sœurs siluriennes peuvent respectivement être, biologiquement parlant, un ordre et un genre. Attribuer aux espèces sœurs la même catégorie pour satisfaire la définition formelle de Hennig peut être logiquement impeccable mais être biologiquement faux. Entre-temps Hennig lui-même a abandonné la prétention de fournir des définitions non arbitraires des catégories supérieures et a arrêté de situer les taxons de rang supérieur dans des catégories (voir ci-dessus). Les objections de Griffiths concernant les critères évolutionnistes de la catégorisation sont fondées sur les définitions cladistiques spéciales de la monophylie et de la phylogénie. Si l’on rejette ces définitions, les objections tombent d’elles-mêmes.
124 On dit parfois que les taxinomistes évolutionnistes ne fournissent pas de définition des catégories. Ce n’est pas vrai. Ce que j’ai souligné, est que la catégorie espèce est la seule catégorie pour laquelle il est possible de donner une définition non arbitraire. Simpson (1961) et moi (Mayr 1969) avons fourni des définitions formelles des catégories supérieures, même si, pour des raisons expliquées plus haut, l’application de ces définitions a ses limites.
125 Ce qui précède sont les critiques exprimées par Griffiths. Plus généralement les cladistes reprochent aux taxinomistes évolutionnistes de ne pas donner de critères simples qui permettent de prendre des décisions en matière de classification. Il me semble que la nature du sujet s’oppose à une approche simpliste. Le nombre de variables que l’on doit considérer lors de la construction d’une classification est si élevé que les méthodes simples ne marcheront pas. C’est la raison pour laquelle le taxinomiste évolutionniste pondère soigneusement les preuves et fait jouer son jugement afin de pouvoir conclure. Il se demande quel rôle un taxon supérieur joue dans l’économie de la nature. Il tient compte de la nature de la percée adaptative qui donne naissance à un taxon (Peters 1972). Bref, il approche avec insistance son matériel en biologiste et en évolutionniste, plutôt que de s’en remettre à des réponses toutes faites. À court terme cela crée certainement des difficultés et des incertitudes, mais qui contesterait le fait qu’une classification qui utilise toute l’information potentiellement accessible est plus informative, plus prédictive, et, de fait, reflète mieux l’évolution passée qu’une classification qui se restreint à la seule information donnée par le pattern de branchement ?
Synopsis
126 Pour être judicieuse, une classification d’un quelconque groupe d’organismes ne peut pas être conçue indépendamment d’une phylogénie bien reconstruite.
127 L’établissement d’un pattern de branchements des diverses lignes phylétiques et la réalisation d’un cladogramme forment une composante d’une telle reconstruction. Hennig a illustré que cela pouvait être réalisé de façon relativement non ambiguë en classant les caractères en caractères apomorphes (dérivés) et plésiomorphes (ancestraux).
128 Le contenu informatif élevé des caractères dérivés (apomorphes) était reconnu par de nombreux taxinomistes bien avant Hennig mais pas suffisamment apprécié et, en fait, entièrement ignoré par certains auteurs. L’insistance mise sur une pondération appropriée des synapomorphies, liée à la théorie cladistique de Hennig, a permis un développement salutaire de la systématique.
129 Une analyse impartiale serait nécessaire si l’on voulait déterminer à quel point les améliorations récentes de la classification des poissons, des insectes, et d’autres groupes sont le résultat d’une application rigoureuse de l’analyse cladistique. Cracraft (1972) a récemment affirmé que l’application de la méthode cladistique allait révolutionner la taxinomie des oiseaux. Néanmoins, après douze pages de discussions, il n’a pas montré un seul cas qui démontrerait que la classification actuellement acceptée est fausse. Tout ce qu’il a été capable de montrer est qu’une classification fondée uniquement sur les points de branchements est parfois différente d’une classification où la divergence phylogénétique a la primeur. Le cas des Sciadoceridae fossiles (voir plus haut) est une autre illustration d’un changement de classification mais pas d’amélioration. Les raisons pour lesquelles la méthode cladistique ne peut pas apporter plus de résultats devraient être évidentes à la lecture des discussions précédentes.
130 Le regroupement cladistique rencontre bien des difficultés même si l’on écarte les décisions inacceptables concernant la catégorisation. Par exemple, il n’y a souvent pas assez de caractères apomorphes, ou bien il y a des doutes sur l’alternative ancestral-dérivé d’un caractère et, finalement, en raison de l’évolution en mosaïque ou parallèle, il peut y avoir des conflits entre les informations fournies par différents caractères apomorphes.
131 Quelle que soit sa valeur, l’analyse cladistique ne donne pas automatiquement une classification. En n’utilisant qu’un seul jeu parmi les deux, correspondant aux données phylogénétiques, la cladistique produit des classifications qui sont moins capables d’être des systèmes de référence généraux que les classifications évolutionnistes car leur contenu informatif est plus pauvre. Les taxinomistes évolutionnistes, parce qu’ils pondèrent l’information provenant des deux sources, génèrent des classifications qui peuvent être critiquées comme étant plus « subjectives », mais celles-ci reflètent plus exactement l’histoire évolutive et sont donc biologiquement plus significatives. Le cladiste croit qu’une simple addition de synapomorphies donnera automatiquement, si l’on peut dire, la classification correcte. Au contraire le taxinomiste évolutionniste a l’impression que seule une pondération soigneuse de toutes les preuves révélera des degrés de parenté significatifs, au sens de relations génétiques inférées. De plus, il a l’impression qu’une classification doit tenir compte des événements adaptatifs majeurs dans l’histoire évolutive, comme devenir terrestre ou aérien, car ils sont plus importants pour la catégorisation des taxons que la seule subdivision des lignes phylétiques. D’autres systématiciens comme Johnson (1970) et Michener (1970 : 20-22) ont récemment présenté la situation en faveur de la systématique évolutionniste (synthétique).
132 Griffiths (1972 : 17) a conclu que si l’on voulait exprimer les effets des degrés de divergences évolutives inégales sur diverses lignes phylétiques, il faudrait construire une classification tout à fait distincte, autrement dit, on devrait avoir plusieurs (au moins deux) jeux de classifications. Cette proposition m’apparaît comme irréalisable. Peters & Gutman (1971 : 256) rejettent aussi l’approche purement formelle de l’école de Hennig et en appellent à une attitude biologique vis-à-vis des données morphologiques ; ce qui inclut une véritable reconnaissance de l’importance des percées adaptatives.
133 Une classification éclectique qui intègre avec un même souci les points de branchement de la phylogénie et les divers aspects de la divergence phylogénétique semble le meilleur chemin vers des classifications biologiquement significatives, permettant le plus grand nombre de larges généralisations. Le conseil de Darwin d’utiliser ces deux sources d’information a été adopté par les meilleurs classificateurs de ces derniers cent ans.
134 En conclusion, il est évident que l’analyse cladistique, quelle que soit son utilité, ne peut pas être transcrite automatiquement en une classification.
Remerciements
La présente version de mon analyse est le résultat de nombreuses révisions, que j’ai pu mener à bien grâce à la coopération généreuse de nombreux amis et correspondants. Les premières versions ont été lues en entier et critiquées par Peter D. Ashlock, Walter Bock, Arthur Cain, Stephen Gould, Otto Kraus, Günther Osche, D.S. Peters et Donn Rosen. Leurs commentaires sont à l’origine d’une réécriture importante qui ajoute, je l’espère, de la clarification. Gareth Nelson, bien qu’en profond désaccord avec la plupart de mes conclusions, m’a grandement aidé en me fournissant des références et des tirés-à-part difficiles à obtenir. J’apprécie profondément sa générosité.Dans un domaine aussi difficile et controversé que la théorie de la classification personne ne peut espérer avoir raison tout le temps. Je serais tout à fait satisfait si mon analyse pouvait servir à clarifier la question.
Références
- Ashlock P., 1971. Monophyly and associated terms. Systematic Zoology 20 : 63-69.
- Ashlock P., 1972. Monophyly again. Systematic Zoology 21 : 430-438.
- Bigelow R., 1956. Monophyletic classification and evolution. Systematic Zoology 5 : 145-146.
- Bock W., 1959. Preadaptation and multiple evolutionary pathways. Evolution 13 : 194-211.
- Bock W., 1963. Evolution and phylogeny in morphologically uniform groups. American Naturalist 97 : 265-285.
- Bock W., 1965. The role of adaptive mechanisms in the origin of higher levels of organization. Systematic Zoology 14 : 272-287.
- Bock W., 1967. The use of adaptive characters in avian classification, in Proceedings of the 14th International Ornithological Congress, Blackwell, Oxford, 61-74.
- Bock W., 1968. Review of Hennig ‘Phylogenetic Systematics’. Evolution 22 : 646-648.
- Bock W., 1969. Comparative morphology in systematics, in Systematic Biology, National Academy of Sciences, Washington, 411-448.
- Bock W. & Miller W., 1959. The scansorial foot of the woodpeckers, with comments on the evolution of perching and climbing feet in birds. American Museum Novitates 1931 : 1-45.
- Brundin L., 1966. Transantarctic relationships and their significance, as evidenced by chironomid midges. With a monograph of the subfamilies Podominae and Aphroteniinae and the austral Heptagyiae. Kungliga Svenska Vetenskapsakademiens Handlingar 4 (11) : 1-472.
- Brundin L., 1972. Evolution, causal biology, and classification. Zoologica Scripta 1 : 107-120.
- Cain A.J, & Harrison G.A., 1960. Phyletic weighting. Proceedings of the zoological Society, London 135 : 1-31.
- Cracraft J., 1972. The relationships of the higher taxa of birds : Problems in phylogenetic reasoning. Condor 74 : 379-392.
- Crompton A.W. & Jenkins F.A. Jr., 1973. Mammals from reptiles : a review of mammalian origins. Annual Review of Earth and Planetary Sciences 1 : 131-155.
- Crowson R.A., 1970. Classification and biology. Heinemann Educ. Books, London.
- Darlington P.J., 1970. A practical criticism of Hennig-Brundin ‘Phylogenetic systematics’ and Antarctic biogeography. Systematic Zoology 19 : 1-18.
- Darwin C., 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. John Murray, London.
- Farris J., 1966. Estimation of conservatism of characters by consistency within biological populations. Evolution 20 : 587-591.
- Farris J., 1967. The meaning of relationship and taxonomic procedure. Systematic Zoology 16 : 44-51.
- Ghiselin M.T., 1969a. The triumph of the Darwinian method. University of California Press, Berkeley.
- Ghiselin M.T., 1969b. The principles and concepts of systematic biology, in Systematic Biology, National Academy of Sciences, Washington, 45-55.
- Ghiselin M.T., & Jaffe L., 1973. Phylogenetic classification in Darwin’s Monograph on the subclass Cirripedia. Systematic Zoology 22 : 132-140.
- Gisin J., 1964. Synthetische Theorie der Systematik. Zeitschrift für zoologische Systematik und Evolutionsforschung 2 : 1-17.
- Greenwood P.H. & Rosen D.E., 1971. Notes on the structure and relationships of the Alepocephaloid fishes. American Museum Novitates 2473 : 1-41.
- Griffiths G.D.C., 1972. The phylogenetic classification of Diptera Cycclorhapha with special reference to the structure of the male postabdomen. Dr. W. Junk N.V. Publishers, The Hague.
- Günther K., 1956. Systematik und Stammesgeschichte der Tiere. Forstschritte der Zoologie 10 : 37-55.
- Günther K., 1971. Abschließende Zusammenfassung der Vorträge und Diskussionen in R. Siewing (dir.), Methoden der Phylogenetik, Erlanger Forschungen Reihe B, 4, 76-87 (voir aussi page 38).
- Heberer G. (ed.), 1943. Die Evolution der Organismen : Ergebnisse und Probleme der Abstammungslehre. Gustav Fischer, Jena.
- Hennig W., 1950. Grundzüge einer Theorie der Phylogenetischen Systematik. Deutscher Zentralverlag, Berlin.
- Hennig W., 1960. Die Dipteran-Fauna von Neuseeland als systematisches und tiergeographisches Problem. Beiträge zur Entomologie 10 : 15-329.
- Hennig W., 1966. Phylogenetic Systematics. University of Illinois Press, Urbana.
- Hennig W., 1969. Die Stammesgeschichte der Insekten. Senckenberg Buch 49, Verlag W. Kramer, Frankfurt/M.
- Hennig W., 1971. Zur Situation der biologischen Systematik, in R. Siewing (dir.), Methoden der Phylogenetik, Erlanger Forschungen Reihe B 4, 7-15.
- Hull D., 1967. Certainty and circularity in evolutionary taxonomy. Evolution 21 : 174-189.
- Hull D., 1970. Contemporary systematic philosophies. Annual Review Ecology and Systematics 1 : 19-54.
- Huxley J. (ed.), 1940. The new systematics. Clarendon Press, Oxford.
- Huxley J., 1942. Evolution : The modern synthesis. Allen and Unwin, London.
- Huxley J., 1958. Evolutionary processes and taxonomy, with special reference to grades. Uppsala Universitets Årsskrift 6 : 21-39.
- Illies J., 1961. Phylogenie und Verbreitungsgeschichte der Ordnung Plecoptera. Verhandlungen der Deutschen Zoologischen Gesellschaft. Zoologischer Anzeiger. Supplementband 25 : 384-394.
- Inger R., 1967. The development of a phylogeny of frogs. Evolution 21 : 369-384.
- Jepsen G., Mayr E. & Simpson G.G., 1949. Genetics, paleontology, and evolution. Princeton University Press, Princeton.
- Johnson L.A.S., 1970. Rainbow’s end : the quest for an optimal taxonomy. Systematic Zoology 19 : 203-239.
- Kiriakoff S.G., 1959. Phylogenetic systematics versus typology. Systematic Zoology 8 : 117-118.
- Kluge A. & Farris J., 1969. Quantitative phyletics and the evolution of Anurans. Systematic Zoology 18 : 1-32.
- Mayr. E., 1942. Systematics and the origin of species. Columbia University Press, New York.
- Mayr E., 1963. Animal species and evolution. The Belknap Press, Harvard University, Cambridge, Mass.
- Mayr E., 1965. Numerical phenetics and taxonomic theory. Systematic Zoology 14 : 73-97.
- Mayr E., 1969. Principles of Systematic Zoology. McGraw-Hill, New York.
- Mayr E. & Short L., 1970. Species taxa of North American birds. Nuttal Ornithological Club, Cambridge, Mass.
- McAlpine J. & Martin J. 1966. Systematics of Sciadoceridae and relatives with descriptions of two new genera and species from Canadian amber and erection of family Ironomyiidae (Diptera: Phoroidae). Canadian Entomologist 98 : 527-544.
- Michener C.D., 1970. Diverse approaches to systematics. Evolutionary Biology 4 : 1-38.
- Mill J. S., 1874. A system of logic, ratiocinative and inductive, being a connected view of the principles of evidence and the methods of scientific investigation. 8th ed., Longsman, Green and Co, London.
- Nelson G., 1971. Cladism as a philosophy of classification. Systematic Zoology 20 : 373-376.
- Nelson G., 1972. Comments on Hennig’s ‘Phylogenetic Systematics’ and its influence on ichthyology. Systematic Zoology 21 : 364-374.
- Osche G., 1961. Aufgaben und Probleme der Systematik am Beispiel der Nematoden. Verhandlungen der Deutschen Zoologischen Gesellschaft 1960 : 329-384.
- Osche G., 1965. Über latente Potenzen und ihre Rolle im Evolutionsgeschehen. Zoologischer Anzeiger 174 : 411-440.
- Osche G., 1971. Discussion comments, in R. Siewing (dir.), Methoden der Phylogenetik, Erlanger Forschungen Reihe B 4, 85.
- Pennak R., 1964. The collegiate dictionary of zoology. Ronald Press, New York.
- Peters D.S., 1970. Über den Zusammenhang von biologischem Artbegriff und phylogenetischer Systematik. Aufsätze und Reden der Senckenbergischen Naturforschenden Gesellschaft 18, W. Kramer & Co, Frankfurt/M.
- Peters D.S., 1972. Das Problem konvergent entstandener Struckturen in der anagenetischen und genealogischen Systematik. Zeitschrift für zoologische Systematik und Evolutionsforschung 10 : 161-173.
- Peters D.S. & Gutmann W.F., 1971. Über die Lesrichtung von Merkmalsund Konstruktions-Reihen. Zeitschrift für zoologische Systematik und Evolutionsforschung 9 : 237-263.
- Popper K.R., 1959. The logic of scientific discovery. Hutchinson, London.
- Popper K.R., 1963. Conjectures and refutations. Routledge and Kegan Paul, London.
- Remane A., 1952. Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. Geest & Portig, Leipzig.
- Rensch B., 1947. Neuere Probleme der Abstammlungslehre. Die transspezifische Evolution. Ferdinand Enke, Stuttgart.
- Schindewolf O.H., 1967. Über den ‘Typus’ in morphologischer und phylogenetischer Biologie. Akademie der Wissenschaften und der Literatur (Mainz), Abhandlungen der Mathematisch-Naturwissenschaftlichen Klasse 4 : 57-131.
- Schlee D., 1971. Die Rekonstruktion der Phylogenese mit Hennigs Prinzip. Aufsätze und Reden der Senckenbergischen Naturforschenden Gesellschaft 20 : 1-62.
- Siewing R. (dir.), 1971. Methoden der Phylogenetik. Erlanger Forshung Reihe B Naturwissentschaften 4, Erlangen-Nürnberg.
- Simpson G.G., 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85 : 1-350.
- Simpson G.G., 1953. The major features of evolution. Columbia University Press, New York.
- Simpson G.G., 1959a. Anatomy and morphology : classification and evolution, 1859 and 1959. Proceedings of the American Philosophical Society 103 : 286-306.
- Simpson G.G., 1959b. The nature and origin of supraspecific taxa. Cold Spring Harbor symposia on quantitative biology 24 : 255-271.
- Simpson G.G., 1961 Principles of animal taxonomy. Columbia Univiversity Press, New York.
- Sokal R. & Sneath P., 1963. Principles of numerical taxonomy. Freeman, San Francisco.
- Throckmorton L., 1962. The problem of phylogeny in the genus Drosophila. Studies in genetics II. University of Texas Publication 6205 : 207-343.
- Throckmorton L., 1965. Silmilarity versus relationship in Drosophila. Systematic Zoology 14 : 221-236.
- Tillyard R., 1921. A new classification of the order Perlaria. Canadian Entomologist 53 : 35-43.
- Warburton F.E., 1967. The purposes of classification. Systematic Zoology 16 : 241-245.