Remarques critiques concernant la question « Analyse cladistique ou classification cladistique ? »
- Par Willi Hennig,
- Traduit par Annemarie Ohler
Pages 71 à 95
Citer cet article
- HENNIG, Willi,
- Traduit par OHLER, Annemarie,
- Hennig, Willi.,
- et al.
- Hennig, W.,
- Traduit par Ohler, A.
https://doi.org/10.3917/biosy.029.0071
Citer cet article
- Hennig, W.,
- Traduit par Ohler, A.
- Hennig, Willi.,
- et al.
- HENNIG, Willi,
- Traduit par OHLER, Annemarie,
https://doi.org/10.3917/biosy.029.0071
Notes
-
[1]
Les répétitions de mots qu’on trouve tout le long de cette traduction sont reprises du texte allemand. Leur effet stylistique est le même en allemand qu’en français. (Ndt.)
-
[2]
Le mot allemand « Stammbaum » s’applique autant aux relations de parenté dans une famille qu’aux relations phylogénétiques entre lignées. Je le traduis comme « arbre phylogénétique », ce qui correspond au « phylogenetic tree » utilisé par Mayr (1974) dans sa critique. (Ndt.)
-
[3]
Nous avons tenté avec ce terme de rendre compte du mot enkaptisch « par emboîtement », utilisé traditionnellement en allemand dans le contexte de la biologie de l’évolution. (Ndt.)
-
[4]
Le terme « adaptiogenetisch » est traduit par « adaptatif ». (Ndt.)
-
[5]
Le mot « aramorphose » du texte allemand doit résulter d’une coquille. (Ndt.)
-
[6]
Hennig écrit littéralement, de façon redondante, « un seul ancêtre commun à eux ». (Ndlr.)
-
[7]
Ici, Hennig utilise le mot anglais « cladogram ». (Ndlr.)
-
[8]
La traduction par Hennig de « ludicrous » (grotesque) par « enorm verwirrende (!) » (énormément déconcertante) est un euphémisme. (Ndlr.)
-
[9]
Hennig utilise ici le mot germanisé « Kladogramm ». (Ndlr.)
-
[10]
Ici Hennig a mal compris le texte de Mayr (p. 34 de la présente traduction) qui indique au contraire que les cladistes ont repris la logique « vers le bas » de la classification d’Aristote au lieu du principe de classification vers le haut qui prédomine depuis Darwin (voir avantpropos, p. 8). (Ndlr.)
-
[11]
En allemand Hennig parle de « konsequentphylogenetisch » et lie le terme phylogenetisch à la manière d’élaborer et d’interpréter les relations phylogénétiques. Je lie ici les deux termes par des guillemets pour indiquer ce lien entre les deux mots. (Ndt.)
-
[12]
En allemand, « vordeszendenztheoretisch » signifie littéralement « pré-descendance-théo-rique », mais le français ne permet pas de créer des termes en accolant ainsi les mots. (Ndt.)
1 Il y a quelque temps déjà, mon collègue Mayr m’a aimablement envoyé son manuscrit Cladistic analysis or cladistic classification ? (« Analyse cladistique ou classification cladistique »). Celui-ci a été publié récemment dans ce journal et contient principalement une critique de la « systématique phylogénétique ». Cependant, de nombreux aspects de cette critique [1] se fondent sur des incompréhensions, des imprécisions logiques et des contradictions, et le tout est alourdi par un traitement détaillé de questions périphériques qui n’ont que peu à voir avec le thème de la critique de Mayr. Par ces aspects, avant tout, la présentation de Mayr donne l’impression de se donner pour but de défendre une pensée véritablement biologique contre une pensée purement formaliste, la pluralité contre l’étroitesse d’esprit et le manque d’expérience, la tradition confirmée contre une manie infondée de renouvellement. Étant donné la grande reconnaissance scientifique dont Ernst Mayr bénéficie, tout cela peut facilement créer la confusion chez quelquesuns. Pour cette raison je veux exceptionnellement déroger à mon principe strictement suivi par ailleurs, de ne pas répondre à la critique par un travail particulier. Cependant je n’ai pas l’intention dans ce qui suit de répondre en détail à toutes les remarques de Mayr qui devraient être contredites. Pour cela mon exposé devrait devenir au moins aussi volumineux que le sien. C’est impossible pour de nombreuses raisons, et peut-être n’est-ce pas nécessaire. Je me borne donc essentiellement à aborder au plus près certaines questions fondamentales.
1.
2 Après des remarques introductives, Mayr (dans le chapitre « La reconstruction du schéma de branchement de la phylogénie – analyse cladistique) » rend pleinement hommage à la légitimité épistémologique et méthodologique et à l’efficacité pratique de la démarche méthodologique de la systématique phylogénétique qui, dans l’évaluation relativisante et dans l’analyse de l’« héterobathmie des caractères », recherche la séquence chronologique des événements de spéciation dans les lignées de descendance phylogénétique, permettant ainsi de reconstruire l’arbre phylogénétique [2] (dans le sens strict d’un cladogramme) (Schlee 1971).
3 Mayr assure ainsi que « l’analyse cladistique » réalisée en appliquant cette méthode est un travail préliminaire indispensable et obligatoire pour chaque taxinomiste, mais il rejette de manière insistante le postulat de la systématique phylogénétique (Mayr : « classification cladistique »), consistant à transformer le cladogramme obtenu en un système hiérarchique. Un tel système, qui tend vers un arrangement emboîté [3] de ces taxons exclusivement selon la séquence chronologique des événements de spéciation dans les lignées phylogénétiques, est, selon les termes de Mayr, « défectueux », « irréaliste » et non scientifique : ce faisant, Mayr expose par la suite une conception d’une évolution phylogénétique parfois adaptative [4] ou écofonctionnelle entièrement divergente de celle des groupes frères phylogénétiques. Comme la systématique phylogénétique classe les groupes frères phylogénétiques dans des catégories de même rang, quand l’un a atteint un niveau d’adaptation tout à fait différent de celui de l’autre qui a conservé le niveau de l’ancêtre commun, selon Mayr le système phylogénétique (dans le sens des « cladistes », comme les appelle Mayr) ne comporte pas d’information sur ce phénomène important de l’adaptation divergente chez les groupes phylogénétiquement frères.
4 Cette conception de la systématique phylogénétique (« cladistique ») critiquée par Mayr n’est seulement concevable selon son affirmation qu’en raison de trois conditions scientifiquement non permises selon lui :
- l’affirmation de la systématique phylogénétique comme étant la meilleure systématique (classification) possible ;
- la « redéfinition arbitraire de termes connus depuis longtemps » comme (a) phylogenèse, (b) parenté et (c) monophylie ;
- la négligence des changements évolutifs adaptatifs divergents dans des lignées phylogénétiques sœurs.
6 À ces objections considérées fondamentales contre la solidité scientifique de la systématique phylogénétique, il sera répondu ci-dessous en détail.
2.
7 Mayr appelle « systématique évolutionniste » la contreproposition qu’il défend. Elle débute également par la construction d’un « cladogramme » mais ne suit pas celui-ci pour la mise en forme d’une classification hiérarchique. En échange, elle considère à la fois des aspects phylogénétiques (« cladistiques ») et des aspects adaptatifs (écofonctionnels). De cette manière elle sauve – selon Mayr – le contenu informatif, qui est également pris en compte par la considération des divergences adaptatives dans l’évolution de groupes frères phylogénétiques. Selon Mayr, la « systématique évolutionniste » répond en même temps – bien plus que le système phylogénétique (« cladistique ») – à l’exigence d’une classification systématique, qui est de permettre, avec sa constitution de groupes, les généralisations les plus larges et les « prédictions » (ainsi que Mayr les appelle) les plus performantes. Mayr qualifie de « théorie » son programme de « systématique évolutive ». Il échange ainsi le concept d’une théorie scientifique avec celui d’un programme de travail arbitraire. Une systématique « évolutionniste » opérant de manière synthétique ou syncrétique selon l’exigence formelle de Mayr ne peut prétendre constituer une théorie scientifique. Cette question a été abordée plusieurs fois, la dernière fois par Johnson (1970), qui lui-même est un représentant de cette systématique synthétique, « évolutionniste ».
8 L’imprécision épistémologique dans les exposés de Mayr est soulignée par le fait qu’il tente d’orienter le système « évolutionniste », explicitement « synthétique », selon deux points de vue différents, un point de vue phylogénétique (cladistique) et un point de vue adaptatif (écofonctionnel). Dans un système reposant sur deux points de vue différents, l’utilisateur d’un groupe (ou taxon) donné ne peut jamais savoir, si ce dernier est construit et défini selon un point de vue phylogénétique (cladistique) ou adaptatif-écofonctionnels. Contrairement à ce qu’écrit Mayr, ceci diminue de manière décisive la valeur informative d’un tel système (voir plus bas).
9 Le vieux postulat – déjà présent chez Aristote, mais pas toujours respecté – de la « stricte unicité du point de vue pour la formation de systèmes », n’est même pas mentionné par Mayr, et encore moins réfuté. Il écarterait cette demande en la qualifiant de « formalisme », comme il le fait par ailleurs pour d’autres nécessités logiques (voir plus bas). Mais ceci n’est pas une réfutation scientifique proprement dite, et la négligence des exigences logiques n’est généralement pas sans inconvénient – également en dehors de la science.
10 Venons-en maintenant au détail des remarques, numérotées plus haut, par lesquelles Mayr essaie de réfuter la solidité scientifique de la systématique phylogénétique.
3.
11 1. « Les cladistes sont sincèrement convaincus que leur théorie produit la meilleure classification. » Cette phrase, avec laquelle Mayr introduit son essai de réfutation de cette revendication supposée, est une erreur, comme le montre sa citation immédiatement après (tirée de Hennig 1971) : je n’ai fait que constater de manière répétitive le primat logique du système (cladistique) phylogénétique (en tant que « système de référence général »), et ce n’est pas pareil à l’allégation de Mayr. La question, quel système classificatoire des animaux est le meilleur, ne peut pas être posée ainsi de manière générale (« le système classificatoire des animaux dans les livres de cuisine est le meilleur pour les utilisations d’un livre de cuisine » : Günther). Mayr donne plusieurs critères pour définir le meilleur système scientifique. Parmi eux, les plus importants et les meilleurs sont « les propriétés explicatives, prédictives et heuristiques » et un « stockage efficace et un système de récupération de l’information ». Ici se pose déjà la question : que signifie le terme « propriété prédictive » dans une théorie de la systématique biologique ? Ne s’agit-il pas ici d’un emprunt purement formel et au fond injustifié auprès d’un groupe particulier de théories scientifiques auquel aucune théorie de la systématique biologique n’appartient ? En dehors de quelques constatations très générales [« Le nombre d’affirmations évolutionnistes et les prédictions qui peuvent être tirées des nombreux groupes holophylétiques (il s’agit des groupes monophylétiques de la systématique phylogénétique) sont souvent minimes »], le terme de « prédiction » ne joue ensuite plus aucun rôle dans l’exposé de Mayr. Il semble que Mayr, contre son intention déclarée, confirme mon opinion du primat logique du système phylogénétique (« cladistique ») : « La méthode synthétique ou évolutionniste de classification est en accord avec la cladistique sur le postulat selon lequel une phylogénie aussi complète que possible doit précéder la construction d’une classification » et (p. 21) « La recherche phylogénétique en tant que science biologique est concevable uniquement si l’on admet que le premier objectif est la découverte de la relation généalogique entre les espèces. »
12 Ici il faudrait signaler une différence qu’on pourrait qualifier comme la distinction entre les termes « système » et « classification ». Ce que cela signifie peut être montré clairement par un exemple : si un archéologue découvre des tessons lors de la fouille d’une tombe, il peut et va (tout d’abord) les ranger (« classifier ») de manière arbitraire : selon le matériau (argile ou métal), selon la couleur, selon le type de décoration, etc. Ensuite il peut aussi essayer de reconstruire à partir des tessons les récipients originels (vases, urnes). Cette activité de reconstruction consiste aussi dans la construction d’un ordre. Mais il est douteux que l’on puisse qualifier l’ordre ainsi obtenu de « classification ».
13 Un autre exemple serait la classification des fleuves d’Europe. On peut les classer selon leur navigabilité, selon leur niveau d’eau, selon les conditions qu’ils offrent pour l’occupation par des organismes, etc. Mais on peut également les étudier en fonction du bassin-versant (du Danube, du Rhin, de l’Elbe) auxquels ils appartiennent.
14 De manière semblable, la construction d’un arbre phylogénétique est une activité de reconstruction et l’ordre des espèces créé par la systématique phylogénétique est quelque chose de fondamentalement différent des autres classifications. En raison d’une intuition (à l’origine encore obscure) en faveur de cette différence, j’ai souvent évité de parler de « classification » phylogénétique (au lieu de « système » phylogénétique), et je ne m’y suis seulement parfois laissé dévoyer que par l’influence de l’usage linguistique anglais.
15 Une étrange erreur dans le travail de Mayr, qui met en doute la valeur de l’ensemble de son exposé supposé fondamental, consiste dans le fait qu’il dénie la convertibilité mutuelle d’un arbre phylogénétique et du système hiérarchique (p. 65) : « Quelle que soit sa valeur, l’analyse cladistique ne donne pas automatiquement une classification », et (p. 25) « le postulat de base de la théorie cladistique, une congruence absolue entre le cladogramme et la classification, ne peut être satisfait qu’en faisant de nombreuses hypothèses et redéfinitions et en ignorant de nombreux faits de l’évolution et de la phylogénie, au sens large ».
16 Cette allégation est purement et simplement fausse. Elle serait seulement juste si, dans les deux phrases, à la place de « classification » on mettait « classification sensu Mayr ». Mais alors nous ne la contredirions pas. Mayr ici confond les intentions de construction des systèmes des « cladistes » avec celles des « évolutionnistes », bien qu’il ait amplement décrit les différences entre les deux, et tout en reprochant aux cladistes de procéder autrement qu’il le souhaiterait. Cette confusion singulière est présentée comme une réfutation des « cladistes ». On la retrouve dans d’autres passages.
17 En réalité la transcription d’un cladogramme (Mayr, Fig. 1) en un système hiérarchique est possible sans une seule supposition ou redéfinition supplémentaire d’un seul des termes employés pour l’ébauche d’une représentation d’arbre phylogénétique. La transformation est un processus purement formel :
Taxon (B + C + D)
- Taxon B
- Taxon (C + D)
- Taxon C
- Taxon D
19 Nous allons développer rapidement pourquoi nous préférons souvent ce mode de notation à une représentation d’arbre phylogénétique.
4.
20 2. Le primat logique du système phylogénétique (cladistique) comme «système de référence général» repose, dans sa fondation, sur une théorie biologique s’appuyant sur des concepts centraux définis rigoureusement, c’est-à-dire univoques. Mayr lui oppose ici l’objection «de redéfinitions arbitraires de termes bien connus». Il en va comme suit :
21 a. «Phylogenèse», un terme façonné il y a plus de 100 ans par Haeckel, n’avait, jusqu’à présent, pas été défini de manière univoque. Le mot grec signifie approximativement «devenir (formation) des phyla = communautés de descendance». Selon la théorie de la descendance, ce processus consiste en des événements de divisions successives de communautés reproductives d’organismes. La systématique phylogénétique (cladistique) définit le terme «phylogenèse» de cette manière univoque. Mayr en revanche considère cela insuffisant et souhaite prendre en compte un deuxième «ensemble de facteurs», la «taille et l’ampleur des changements évolutifs entre les événements de division». Pour ces épiphénomènes de la phylogenèse, plus ou moins importants, qui apparaissent parfois chez des groupes frères, nous avons les termes adaptation («adaptogenèse») et anagenèse (anamorphose [5]), qui sont utilisés dans la littérature avec différentes significations. Peut-être cela provient-il d’un sentiment qu’il s’agit là d’épiphénomènes logiquement subordonnés à la phylogenèse, dans la mesure où Mayr appelle la démarche classificatoire préconisée par lui comme étant évolutionniste et non pas comme classification (proprement) phylogénétique.
22 b. «Parenté» (relationship). Ici aussi la critique de Mayr se fonde de manière évidente sur un manque de compréhension de la nécessité de termes définis de manière non ambiguë. En langue allemande comme en anglais, «parenté» signifie aussi bien «parenté de forme» que parenté généalogique (de «sang»). Mayr défend cette ambiguïté du terme «parenté» (c’est-à-dire relationship) dans le cadre de la méthode de la systématique biologique, sans remarquer qu’il défend ainsi une logique impossible ; car on ne peut pas travailler de manière scientifique avec des termes ambigus et non définis pour une application spécifique. Pour cela la systématique phylogénétique définit le terme «parenté» rigoureusement et univoquement comme parenté «généalogique (de sang)». «Rigoureusement et univoque» sont rejetés par Mayr comme «hyperspécialisés», ce qui qualifie également son argumentation d’irrecevable.
23 c. « Monophylie». La manière dont Mayr, dans son rejet de définitions rigoureuses et univoques pour des termes scientifiques qui n’ont jamais été définis de manière logique, crée de la confusion au lieu de la clarté, est illustrée dans son exposé concernant le concept de monophylie.
24 Mayr défend la définition «traditionnelle» du terme et soutient (p. 32) que j’aurais créé une énorme confusion par la limitation du sens du terme. En vérité la définition traditionnelle du terme monophylie, comme l’entend Mayr («descendance à partir d’un ancêtre commun»), est absolument vide de sens. Si Mayr (p. 17) pense que la concordance entre systématique phylogénétique («cladistique») et systématique évolutionniste, réside dans le fait que «les groupes qui ne sont pas composés de descendants d’un ancêtre commun sont artificiels et ont une valeur prédictive faible», cela est vide de sens, parce que chaque paire quelconque d’espèces animales possède un ancêtre commun. Selon ce critère, n’importe quelle formation de groupe serait permise. C’est seulement grâce à l’assertion que des espèces animales doivent partager un ancêtre commun exclusif [6] pour pouvoir être conçues comme membres d’un groupe monophylétique, que la définition devient claire et utilisable.
25 Dans un certain sens, l’indétermination de l’énoncé, selon lequel des groupes monophylétiques doivent avoir «un ancêtre commun», dans la définition traditionnelle comme la comprend Mayr (p. 31), est restreinte par le complément «en raison de leurs caractéristiques, les espèces qui composent le groupe sont les plus proches parentes les unes des autres». Toutefois la valeur de cette formulation est annulée par l’indétermination du terme «plus proches parents», si on ne comprend pas, par cela, les parents au sens généalogique du terme. Ce faisant, seule persiste une différence de précision dans la formulation entre la «définition» traditionnelle du terme de monophylie avec ses deux composants mentionnés par Mayr et la définition de la systématique phylogénétique.
26 Effectivement il est caractéristique, dans la situation présente de la systématique, que beaucoup de systématiciens souhaitent voir élargir le sens du terme «monophylétique» pour y inclure également la formation de groupes que j’ai appelés «paraphylétiques». Mayr cite ici Ashlock qui introduit la désignation «holophylétique» pour ces groupes qui peuvent être considérés comme monophylétiques dans le sens de ma définition (parce qu’ils n’ont qu’un seul ancêtre commun qui leur est propre), et veut unir ces groupes holophylétiques avec les paraphylétiques sous la dénomination commune de «monophylétique». On rencontre des groupes typiquement paraphylétiques dans une classification qui oppose l’homme «aux animaux», et dans une classification qui oppose invertébrés et vertébrés. Si l’on désignait alors ces groupes paraphylétiques («animaux», «invertébrés») comme monophylétiques, comme cela résulte de la définition d’Ashlock, alors ce terme perdrait toute valeur pratique.
27 Par ailleurs l’essai d’Ashlock, d’unir sur la base de certaines concordances et différences supposées dans la structure des relations généalogiques, des groupes «paraphylétiques» et monophylétiques (holophylétiques) (les groupes monophylétiques d’Ashlock) et de les opposer aux groupes polyphylétiques, est condamné définitivement à l’échec pour des raisons théoriques. Une différence catégorique existe en vérité entre les groupes monophylétiques (dans le sens de la définition défendue par moi ; = groupes holophylétiques dans le sens d’Ashlock), qui n’ont qu’une seule espèce ancestrale exclusive et des groupements non monophylétiques (paraphylétiques et polyphylétiques), dont les composants ont également toujours une espèce ancestrale commune, mais aucune seulement commune à eux.
28 Une distinction terminologique entre groupes paraphylétiques et polyphylétiques n’est rendue possible que par des moyens méthodologiques. Si l’on découvre, en analysant de manière critique un regroupement indiscutablement monophylétique, que la classification acceptée jusqu’à maintenant a distingué dans son sein deux sous-groupes non monophylétiques, il s’avère toujours qu’une des deux erreurs possibles a été faite (Fig. 1). Les sous-groupes ont soit été formés par des concordances entre leurs composants qui se sont ensuite avérées comme des convergences. Des tels groupes sont désignés depuis longtemps comme «polyphylétiques». Les sous-groupes peuvent aussi avoir été formés pour des raisons de symplésiomorphie, et on a pu démontrer que les espèces réunies en leur sein n’ont pas d’ancêtre commun exclusif. Ces groupes ont été désignés dans la littérature parfois comme polyphylétiques, parfois comme monophylétiques. Comme le montre la figure, aucune différence n’existe dans la structure des relations généalogiques entre les espèces de tels groupes, ceux que j’ai désignés comme paraphylétiques, et les groupes polyphylétiques. La différenciation terminologique entre les groupes paraphylétiques et polyphylétiques est ainsi seulement justifiée si l’on veut souligner le type particulier d’erreur dans l’analyse des caractères qui a mené à la formation de ces groupes. Pour la description des différences dans les relations généalogiques de leurs composants, les désignations «groupe paraphylétique» et «groupe polyphylétique» ne peuvent pas être utilisées. Dans le contexte de la confusion qui a été créée par Ashlock et l’utilisation de sa terminologie par Mayr et Weidner, il n’y a pas d’autre issue dans l’intérêt de la précision terminologique que d’abandonner entièrement le terme ambigu «monophylétique», ou bien dans le futur de ne l’utiliser que dans le sens défini par moi. Dans la période transitoire on pourrait par exemple parler de «groupes monophylétiques (holophylétiques)».
Schéma explicatif des groupes polyphylétiques et paraphylétiques. Les espèces A, C et E s’accordent en fonction de caractères ancesteraux (plésiomorphes) cercles blancs), les espèces B, D et F s’accordent en fonction de caractères dérivés mais dus à la convergence. Voir également le texte.
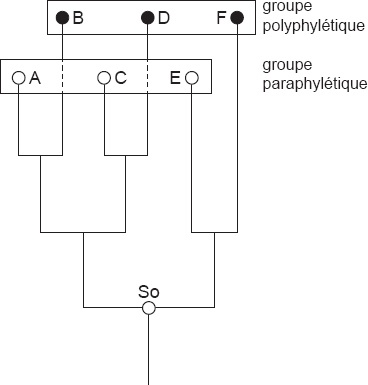
Schéma explicatif des groupes polyphylétiques et paraphylétiques. Les espèces A, C et E s’accordent en fonction de caractères ancesteraux (plésiomorphes) cercles blancs), les espèces B, D et F s’accordent en fonction de caractères dérivés mais dus à la convergence. Voir également le texte.
29 Contrairement à l’affirmation de Mayr, la définition du terme «monophylétique» ne devient pas importante seulement en transposant un arbre phylogénétique en un système hiérarchique, mais déjà lors la construction de l’arbre phylogénétique (analyse cladistique, mise en évidence d’une séquence chronologique des points de branchements de l’arbre phylogénétique, degré d’ancienneté de l’ascendance commune ou ancestralité). Un «cladogramme» [7], qui ne montre pas exclusivement les groupes monophylétiques (holophylétiques) n’est pas un arbre phylogénétique à notre sens. Si Mayr ne voit pas cela, toutes ses concessions faites aux valeurs de «l’analyse cladistique» et son affirmation selon laquelle «Il n’y a guère d’opposition entre taxinomistes cladistes et évolutionnistes à propos des cladogrammes qui résultent de l’analyse cladistique» deviennent également caduques.
5.
30 3. La négligence du caractère dualiste des modifications évolutives. Mayr regrette que ma définition univoque «énormément déconcertante» (!) [8] du terme monophylie utilisée dans le système «méconnaît et même manque de manière entièrement délibérée l’aspect le plus intéressant de l’évolution et de la phylogenèse», à savoir celui de l’adaptation radiative et divergente. Avec cette définition, la systématique phylogénétique est forcée, par exemple, à former le groupe monophylétique des Archosauromorpha, dans lequel les crocodiles sont unis avec les oiseaux. Mayr qualifie d’«inutile» ce regroupement; mais n’est-ce pas justement l’information considérée manquante par Mayr dans le système phylogénétique (cladistique) sur les divergences adaptatives dans les groupes frères phylogénétiques, «la double nature du changement évolutif» (Mayr), avec un groupement des crocodiles et des oiseaux dans un taxon, qui apparaît de manière plus rigoureuse que dans les classifications habituelles qui ne laissent pas du tout apparaître la relation de groupes frères phylogénétiques entre les crocodiles et les oiseaux :
32 En fait, Mayr devrait, avec son critère de la «négligence de la double nature du changement évolutif», rejeter aussi la formation de groupes comme celui des chordés, dans lequel ont eu lieu des adaptations aussi divergentes que celles des Tuniciers et des Aves, ou bien celui des Mammalia, qui a réalisé des adaptations aussi divergentes que celle des Monotremata et des Proboscidea. Les réflexions dans cette direction, qui pourraient être élargies sans limite, montrent cependant que la question de savoir comment et dans quelle mesure les représentations molles et vagues de Mayr pourraient devenir effectives de manière scientifique et univoque, ne sera jamais clarifiée.
33 Il ne reste alors rien de plus des arguments de Mayr que l’affirmation non fondée (p. 63) : «Attribuer aux espèces sœurs la même catégorie […] peut être logiquement impeccable mais être biologiquement faux»; ici comme ailleurs «dans sa polémique contre la systématique phylogénétique, Mayr se place au-dessous de son niveau» (Löther 1972 : 237).
6.
34 La systématique (plus exactement «classification») évolutionniste défendue par Mayr se sert également du système hiérarchique. Selon ses indications, elle considère ce faisant, en dehors de la séquence des branchements de l’arbre phylogénétique, également un second «ensemble de facteurs» (ou ailleurs «ensemble de variables»), à savoir «la quantité et la nature du changement évolutif entre les points de branchement». Il en résulterait que le système de la «systématique évolutive» aurait un contenu informatif bien plus important que le système de la systématique phylogénétique. Il nous faut maintenant examiner si cela est vrai. L’arbre phylogénétique des Sauropsides récents ressemble, selon le consensus général, à la figure qui suit (Fig. 2).
35 Traduit dans un schéma hiérarchique dans lequel les relations phylogénétiques des Testudines restent ouvertes :
- Tortues
- Archosauroporpha (Huene)
- Crocodylia
- Aves
- Lepidosauria
- Rhynchocephalia
- Squamata
Arbre phylogénétique des Sauropsida selon l’acceptation générale d’auteurs récents. Les relations de parenté des Testudines sont non élucidées. Voir également le texte et la Fig. 3.

Arbre phylogénétique des Sauropsida selon l’acceptation générale d’auteurs récents. Les relations de parenté des Testudines sont non élucidées. Voir également le texte et la Fig. 3.
37 Le contenu informatif de ce schéma consiste en ceci qu’il reflète de manière absolument fiable la séquence des points de branchements tels qu’ils sont connus. On peut les reconnaître aussi clairement que dans un arbre phylogénétique (cladogramme [9]). Cela montre aussi que les relations de parenté des tortues sont inconnues. S’il s’avérait que les tortues sont le groupe frère phylogénétique des autres Sauropsida, la notation devrait se présenter ainsi :
- Tortues
- (par exemple : Sauropsida s. str.)
- Lepidosauria
- Rhyncocephalia (sic)
- Squamata
- Archosauromorpha
- Crocodylia
- Aves
39 Le système des Sauropsides, qui correspondrait à la «classification évolutive», ressemble à ceci :
Classe Reptilia
Classe Aves
- 1. Ordre Chelonia (Testudines)
- 2. – Rhynchocephalia
- 3. – Squamata
- 4. – Crocodylia
41 Si l’on voulait transposer ce schéma dans un cladogramme, on obtiendrait l’«arbre phylogénétique» suivant (Fig. 3).
Le «cladogramme», qui résulterait de l’essai de reconstruction d’un «arbre phylogénétique» à partir de la division habituelle («évolutionniste») des Sauropsida en classes et ordres. Voir le texte et la Fig.2.

Le «cladogramme», qui résulterait de l’essai de reconstruction d’un «arbre phylogénétique» à partir de la division habituelle («évolutionniste») des Sauropsida en classes et ordres. Voir le texte et la Fig.2.
42 Cet «arbre phylogénétique» serait cependant entièrement faux. Le système évolutionniste ne fournit donc ici aucune donnée sur la séquence de branchements de l’arbre phylogénétique, alors qu’elle devrait être considérée dans le système (le «premier ensemble de facteurs»). Dans d’autres cas, l’arbre phylogénétique transcrit sans information complémentaire selon le même principe d’un système évolutionniste, donne la séquence des branchements de manière partiellement juste. Il montre dans certaines parties effectivement la séquence juste des points de branchements. Mais sans information complémentaire il n’est pas possible de reconnaître dans quelles sections c’est le cas.
43 Si maintenant le «système évolutionniste» ne contient pas d’information fiable sur le premier «ensemble de facteurs» qui a été prétendument inclus dans le système, comment se comporte la fiabilité de l’information sur le deuxième jeu de facteurs «la nature et la quantité du changement évolutif entre les points de branchement»? Ce deuxième jeu de facteurs est, selon la présentation de Mayr, extraordinairement complexe. Il inclut également, par exemple, (p. 64) le «rôle [qu’]un taxon supérieur joue dans l’économie de la nature».
44 Il faudrait ici de nouveau poser la question de savoir quels sont les critères objectifs et obligatoires pour chaque systématicien permettant de transformer quantitativement ces nombreuses choses fortement différentes dans des valeurs quantitatives (parce qu’il s’agit de cela finalement) pour en définir la coordination ou la subordination des groupes dans le système.
45 Un tel critère ne figure nulle part dans la littérature, y compris dans les travaux de Mayr, et il n’y a pas non plus ici les prémisses d’une méthode praticable qu’on pourrait enseigner. Sur ce sujet il existe une littérature abondante et je ne peux pas l’aborder plus amplement ici.
46 Une autre chose encore est importante dans ce contexte et peut-être plus importante que toute autre. Si l’on pose la question : comment s’expriment effectivement la «nature et la quantité de changement évolutif», la réponse ne peut être que «la nature et la quantité» des multiples différences (les plus diverses) qui existent entre des organismes. Si l’on veut inclure la nature et la quantité des différences en tant qu’information dans un système hiérarchique, il ne s’agit effectivement pas d’informations sur le processus de la phylogenèse ou de l’évolution en soi, mais sur un résultat de ce processus (voir plus haut : l’adaptation comme épiphénomène de la phylogenèse). Si l’on veut classer ce résultat, il est largement indifférent de savoir s’il a été engendré par la phylogenèse (ou évolution) ou si l’on suppose que cela s’est fait par un acte de création unique. Mayr doit avoir senti cela quand il se réfère de manière répétée à Aristote et quand il reproche à la systématique phylogénétique de s’être éloignée de «la classification vers le bas d’Aristote» [10]. En fait si Aristote, en essayant de classer les organismes selon «nature et quantité» des différences avait été doté de nos connaissances contemporaines sur les différences (au sens le plus large) entre les organismes, mais sans connaître les mécanismes de l’évolution et de la phylogenèse, il aurait abouti au même résultat que les adeptes de la manière de classer préconisée par Mayr. Excepté seulement que lui non plus n’aurait pas eu d’échelle obligatoire, objective, pour la mesure des différences morphologiques («Gestaltveränderung»). La dénomination «systématique évolutionniste» est en conséquence trompeuse et bien moins justifiée que systématique «aristotélicienne» (Crowson) ou «typologique».
47 Les discussions concernant les mérites divers de la systématique phylogénétique et de la manière de classer aristotélicienne (Crowson 1970) («systématique évolutionniste» au sens de Mayr) sont véritablement perturbées par deux aspects qui ont entraîné une certaine confusion.
48 Le premier consiste en ceci que les principes de la systématique phylogénétique sont souvent expliqués par l’exemple des Amniotes ou des Sauropsides. C’est commode parce que chacun relie aux noms des groupes de vertébrés une image parlante, mais cela induit certaines difficultés : la conclusion que, dans un système phylogénétique, le groupe «Reptilia» devrait être dissous, et que Crocodylia et oiseaux devraient être unis dans un groupe (Archosauromorpha), agit de manière choquante et «absurde» sur de nombreuses personnes parce que «la nature et la quantité du changement évolutif» (ou bien «l’ampleur des étapes anagénétiques» comme d’autres l’ont formulé) semblent séparer les oiseaux de manière si évidente de tous les «reptiles» (incluant les Crocodylia) qu’ignorer ce fait dans une classification semble être un pur formalisme et un perfectionnisme dépassant tout but raisonnable. On pourrait aussi démontrer ici que le groupement «Archosauromorpha» n’est pas tout à fait «inutile» comme le soutient Mayr. Mais je veux me satisfaire de l’indication qu’il serait trop facile d’argumenter avec un seul cas extrême.
49 Dans les Fig. 4 A-E, plusieurs systèmes sont présentés, qui sont défendus à peu près en même temps dans différents manuels sur les insectes. Ils se ressemblent dans l’intention de représenter dans la classification la différence morphologique des groupements, «la nature et la quantité de changement évolutif» ou «l’ampleur des étapes anagénétiques». Leur différence consiste en ce que ces critères de classification sont évalués de manière tout à fait différente par les différents auteurs (voir les longueurs des flèches).
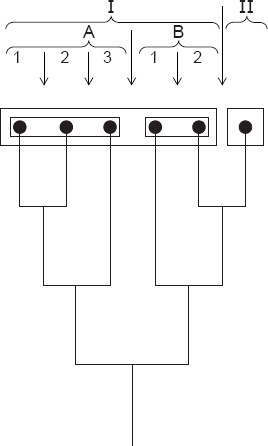
Système «strictement phylogénétique» des Insectes. La division hiérarchique des sous-groupes résulte ici des séquences chronologiques des pas de branchement de l’arbre phylogénétique. Cet arbre phylogénétique peut être reconstruit de manière exacte à partir du système hiérarchique.

Système «strictement phylogénétique» des Insectes. La division hiérarchique des sous-groupes résulte ici des séquences chronologiques des pas de branchement de l’arbre phylogénétique. Cet arbre phylogénétique peut être reconstruit de manière exacte à partir du système hiérarchique.
A. EntognathaI. Ellipura
1. Protura
2. Collembola
II. Diplura
B. Ectognatha
I. Archaeognatha
II. Dicondylia
1. Zygentoma
2. Pterygota
Classification des Insectes (d’après Grassé 1949, Traité de zoologie).
Classification des Insectes (d’après Grassé 1949, Traité de zoologie).
I. ApterygotaA. Entotropha
1. Protura
2. Collembola
3. Diplura
B. Ectotropha
1. Archaeognatha
2. Zygentoma
II. Pterygota
50 Il faut relever ici que le système figuré en Fig. 4 E n’est pas faux dans le sens de la systématique phylogénétique, mais seulement incomplet : il omet plusieurs des données disponibles sur les relations phylogénétiques de certains groupes monophylétiques.
51 On peut montrer des exemples semblables pour la plupart des groupes animaux. Aussi longtemps qu’aucune échelle obligatoire et objective ne peut être indiquée, à l’aide de laquelle on pourrait mesurer «la nature et la quantité de changement évolutif» ou «l’ampleur des étapes anagénétiques», ce qui permettrait de démontrer qu’une des classifications données est plus juste que les autres, il ne peut exister et il n’existera pas de classification aristotélicienne («systématique évolutionniste») généralement acceptée.
Classification des Insectes (d’après Weber 1954,Grundriß der Insektenkunde). 1-5 : «Sous-classes».

Classification des Insectes (d’après Weber 1954,Grundriß der Insektenkunde). 1-5 : «Sous-classes».
1. Collembola2. Protura
3. Diplura
4. Thysanura
5. Pterygota
Classification des Insectes (d’après Beier 1969,Handbuch der Zoologie). sC : sous-classe ; SO : super-ordre ; O : ordre ; sO : sous-ordre.

Classification des Insectes (d’après Beier 1969,Handbuch der Zoologie). sC : sous-classe ; SO : super-ordre ; O : ordre ; sO : sous-ordre.
I. sC. EntergnathaA. SO. Collembola
B. SO. Diplura
1. O. Diplura (sic)
2. O. Protura
II. sC. Ectognatha
(seule O. : Thysanura)
1. sO. Machilinea
2. sO. Lepismatinea
III. sC. Pterygota
Classification des Insectes (d’après Renner 1971, Zoologisches Praktikum). a et b : groupes sans désignation de rang ; sC : sous-classe.
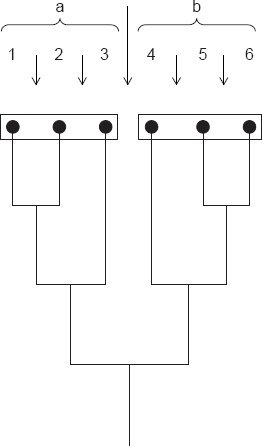
Classification des Insectes (d’après Renner 1971, Zoologisches Praktikum). a et b : groupes sans désignation de rang ; sC : sous-classe.
a. Entotropha1. sC. Diplura
2. sC. Protura
3. sC. Collembola
b. Ectotropha
4. sC. Archaeognatha
5. sC. Zygentoma
6. sC. Pterygota
52 Quand il y a dans l’histoire de la systématique un certain progrès en direction d’un système généralement accepté au moins dans certains groupes animaux, le progrès consiste dans le rapprochement progressif vers un système «strictement phylogénétique» [11].
53 Le second aspect créant de la confusion se laisse aussi reconnaître dans les Fig. 4 AE. Il consiste en ceci que dans les classifications aristotéliciennes comme dans le système phylogénétique les limites des groupes («taxons») coïncident toujours avec les points de branchements des arbres phylogénétiques. Il ne faut que peu de réflexion pour comprendre que ceci doit être le cas et pourquoi. Pour cette raison on peut aussi constater, dans chaque classification élaborée avant l’époque de la théorie de la descendance [12], que les limites des groupes coïncident toujours avec les points de branchement correspondants de l’arbre phylogénétique, pour autant que nous le connaissions aujourd’hui. La différence entre une classification élaborée avant la période de la théorie de la descendance et une autre formée selon les critères recommandés par Mayr consiste seulement dans notre savoir selon lequel les différences entre les différents groupes animaux qui apparaissent plus ou moins importantes à nos yeux, se sont formées au cours du processus phylogénétique (en relation avec la spéciation). En revanche, ce savoir complémentaire (explicatif) est ensuite sans aucune signification pour la théorie et les méthodes de la systématique biologique, si l’on suit les principes recommandés par Mayr : la différence entre les oiseaux et les crocodiles d’un côté et les crocodiles et les autres groupes de «reptiles» reste la même (aussi grand ou petit, aussi signifiant ou insignifiant que soit l’aspect pris en considération), si on la considère sans explication, qu’elle soit le résultat d’un acte de création unique ou résulte du fait qu’au cours de la phylogenèse différents points de branchement de l’arbre phylogénétique se sont avérés diversement significatifs relativement à la formation de la discontinuité actuelle dans la diversité de formes et de vie des organismes. Ceci nous ramène à la constatation que la désignation «systématique évolutionniste» est trompeuse et qu’il n’existe pas de différence fondamentale entre celle-ci et la méthode de classification d’avant l’époque de la théorie de la descendance, aristotélicienne.
54 Avec la démonstration (voir plus haut Fig. 1) qu’il n’existe pas de différence par rapport aux relations généalogiques des parties entre groupes para- et polyphylétiques, tombe aussi l’argument selon lequel la classification évolutionniste se distingue de celle précédant la période de la théorie de descendance par l’exclusion des groupes polyphylétiques.
7.
55 Jusqu’ici seuls ont été considérés des systèmes qui sont présentés sous la forme sèche d’une check-list. D’autres informations peuvent bien entendu être ajoutées sous forme de textes, autant dans le système phylogénétique que dans une classification aristotélicienne. Cela se pratique au moins pour des données concernant la distribution même dans des check-lists. Mais le système phylogénétique se laisse bien plus facilement compléter par une riche source de données fiables que le système aristotélicien, qui est déjà déterminé dans sa structure par une combinaison opaque de points de vue qualitativement divers. J’invite ici simplement à réfléchir à cet aspect.
56 Contre la systématique phylogénétique, Mayr souligne que «souvent le nombre de caractères disponibles est insuffisant». Ceci n’est que trop juste. Mais ceci constitue également une stimulation énorme pour des recherches futures, et souvent des lacunes évidentes dans le système phylogénétique ont mené à des recherches ciblées qui ont ensuite été couronnées d’un grand succès (Schlee, Zwick entre autres).
57 La raison pour laquelle nous ne nous satisfaisons pas de la représentation des résultats des recherches sur les séquences de branchements dans l’arbre phylogénétique d’un groupe animal par le cladogramme dans le sens de Mayr, mais préférons la forme de représentation hiérarchique, peut être clarifiée par une dernière réflexion.
58 La forme de représentation hiérarchique a quelques avantages. Elle peut être donnée clairement dans un espace relativement étroit. Pour les oiseaux par exemple, elle prendrait le même volume que la check-list de Wetmore. Elle permettrait aussi de rendre évidentes pour chacun, rapidement et clairement, les lacunes des connaissances si, par exemple, les espèces d’un genre monophylétique dont les relations de parenté sont inconnues, sont présentées par ordre alphabétique ou si des groupes dont la monophylie est douteuse sont mis entre parenthèses ou indiqués par des points d’interrogations. Il s’agit là d’une question purement technique. Il faut prendre la mesure de la valeur d’un tel système présenté dans la forme simple d’une check-list comme source d’information et comme stimulation pour le comblement des lacunes dans notre connaissance de la séquence des points de division de l’arbre phylogénétique. Est-ce qu’un système traditionnel ou une classification aristotélicienne construite selon les recommandations de Mayr peut vraiment avoir la même valeur comme stockage d’information et comme source de stimulation pour des recherches nouvelles ? Je crains qu’un catalogue des 300 000 espèces de coléoptères, même construit rigoureusement selon les principes de la classification aristotélicienne recommandée par Mayr, n’intéresse que le spécialiste des coléoptères.
8.
59 En dehors de sa critique manifestement infondée des questions fondamentales discutées ci-dessus, avec laquelle de manière incompréhensible Mayr croit réfuter la systématique phylogénétique («cladistique»), cet auteur a ensuite encore traité longuement quelques questions subsidiaires à son sujet dans une approche largement critique ; seuls quelques aspects seront abordés ici. Il me reproche une «une définition de l’espèce purement formelle» en se référant à une remarque sur la règle de déviation, qui a été reproduite par Schlee (1971 : 28).
60 Jusqu’ici j’ai considéré que le «concept biologique d’espèce» utilisé par moi (depuis 1950) (dans l’essentiel : espèce = communauté de reproduction) ne diffère pas de manière essentielle de celui utilisé par Mayr (Hennig 1950, 1960). Il s’appuie certes sur l’étude d’espèces récentes. Mais si l’on considère que malgré toutes les difficultés et contraintes dans le détail (dont nous sommes bien conscients), les espèces récentes, qui sont indiquées dans chaque check-list complète, correspondent, pour l’essentiel, au concept d’espèce biologique, ceci devrait également correspondre aux espèces qu’on pourrait par exemple faire figurer sur une check-list des espèces du Miocène. Naturellement il est pratiquement impossible d’établir une telle check-list complète. Mais il faut accepter cela comme un scénario possible. Entre les espèces du Miocène et les espèces actuelles d’un groupe monophylétique, sont donc envisageable trois et seulement trois possibilités de relations généalogiques : 1) quelques espèces du Miocène n’ont pas de descendants actuels, 2) une certaine espèce récente se compose des descendants (dans le présent) d’une espèce du Miocène, 3) les descendants encore vivant d’une espèce du Miocène forment aujourd’hui plusieurs espèces. Dans le cas mentionné en 2), il pourrait se faire que les descendants récents de l’espèce du Miocène ne se distinguent pas, dans leurs caractères visibles, des représentants fossiles ou bien s’en distinguent beaucoup. Dans l’un ou l’autre cas ceux-ci ne nous indiqueront pas s’ils appartiennent à une même espèce ou à plusieurs, dans le sens du concept d’espèce biologique. Si, dans un certain cadre, seules les relations généalogiques nous intéressent, ce fait est négligeable, parce que dans chaque cas il s’agit du même taxon, indépendamment du fait que l’espèce du Miocène et ses descendants récents appartiennent ou non à la même (potentielle) communauté reproductive et sont ainsi conspécifiques ou non selon le concept d’espèce biologique. Comme on sait maintenant que déjà des sous-populations d’espèces récentes, qui se sont formées seulement depuis un temps relativement court, se distinguent peu entre elles, et comme chaque entomologiste sait que les différences morphologiques de différentes espèces récentes sont souvent difficiles à trouver (Drosophila, Phlebotomus), on peut bien considérer que, dans la plupart des cas, sinon toujours, les descendants récents d’une certaine espèce du passé géologique ne vont se distinguer de celle-ci que peu, indépendamment du fait qu’elles forment aujourd’hui une ou plusieurs espèces. La règle de déviation et la figure explicative dont il s’agit ici (Hennig 1950 : Fig. 25) ne sont alors pas fausses dans ce sens. La figure chez Schlee, violemment critiquée par Mayr, n’a pas d’autre sens que de montrer ceci. Non seulement la critique de Mayr, mais également les explications de Peters pour lesquels il manifeste une forte approbation, passent totalement à côté du sujet.
9.
61 Mayr se préoccupe ensuite de savoir si, dans la division d’une espèce en désormais deux espèces, on ne devrait pas parler plutôt de la continuité de l’«espèce mère a» à côté d’une «espèce fille b», tandis que la systématique phylogénétique préfère dire ici : l’espèce a cessé d’exister ; nous avons affaire après la division de l’espèce souche à des espèces filles b et c. Mayr ne remarque pas que cela reste une simple dispute autour des mots (ni lui ni personne d’autre ne connaît le pool génique des espèces mères et filles avant et après la division). Ainsi il s’agit simplement de la question concernant la description du processus et du résultat de la spéciation. Celle des «cladistes» est méthodiquement la meilleure (Günther 1962 : 279).
62 Mayr souhaite enfin dans un grand effort invalider le «principe cladistique de la dichotomie», parce que les divisions phylogénétiques se dérouleraient aussi de manière multiple et radiative.
63 Entre-temps, il a déjà été démontré par d’autres auteurs dans le but de contredire cette objection relevée aussi par Darlington, qu’également dans le cas où une division strictement dichotomique de l’arbre phylogénétique ne peut pas être prouvée, il ne doit jamais être conclu qu’il existe en effet une spéciation simultanée multiple. Mayr ne considère pas ceci. D’ailleurs la supposition (empirique et également injustifiée pour des raisons strictement théoriques) qu’il existe pour chaque groupe monophylétique un seul groupe frère, a une valeur heuristique importante : elle contient l’invitation à vérifier, partout où cela ne semble pas le cas et où aucune dichotomie ne semble démontrable, par des recherches particulièrement soigneuses, si le groupe frère ne peut pas être découvert.
10.
64 La section «Le mode d’origine des taxons supérieurs» se fonde de nouveau sur la confusion entre «l’approche cladistique» et celle de la «systématique évolutionniste» et inspire à Mayr la conclusion d’opposer, à tort, aux «cladistes» l’«arbre phylogénétique» que Throckmorton (1965) a conçu pour les Drosophilinae (la Fig. 5 de Mayr). Cet arbre a été construit selon les principes des évolutionnistes et est ainsi le témoin du constat fait plus haut (p. 83) : un dessin ressemblant à un arbre phylogénétique, mais pour la construction duquel il n’a pas été fait appel à des termes (monophylie, parenté) définis au sens de la systématique phylogénétique, ne peut pas être considéré comme un arbre phylogénétique. Ainsi l’argumentation de Mayr reste un nonsens épistémologique. D’ailleurs, la conséquence de son argumentation, est que l’espèce Drosophila tripunctata mise au bout de cet «arbre phylogénétique» devrait être l’«espèce mère» de tous les autres taxons qui y figurent, ce que ni Mayr ni Throckmorton n’iraient prétendre.
65 Les chapitres restants de Mayr, qui concernent des questions isolées du modus operandi des «cladistes», verraient beaucoup de leurs doutes résolus par l’étude de travaux particuliers de différents praticiens de la systématique phylogénétique. Les apories occasionnelles, dont les occurrences dans la systématique phylogénétique sont soulignées si emphatiquement par Mayr, ne peuvent pas non plus être évitées dans sa «classification évolutionniste». La critique de la systématique phylogénétique par Mayr me semble en conséquence infondée. Ses incontestables grands mérites se situent ailleurs.
66 Il était très important pour moi de m’assurer que je n’ai ni mal compris ni critiqué de manière injustifiée l’exposé d’Ernst Mayr. Pour cela je suis très reconnaissant au Professeur Dr. K. Günther, Berlin, d’avoir lu soigneusement le projet de mes réponses, et de l’avoir critiqué et complété sur des points essentiels. Je dois la même reconnaissance pour son aide au Dr. D. Schlee, Ludwigsburg.
Références
- Crowson R.A., 1970. Classification and Biology. Heinemann Educ. Books, London.
- Günther K., 1962. Systematik und Stammesgeschichte der Tiere 1954-1959. Fortschritte der Zoologie 14 : 268-547.
- Hennig W. 1950. Grundzüge einer Theorie der Phylogenetischen Systematik. Deutscher Zentralverlag, Berlin.
- Hennig W., 1966. Phylogenetic Systematics. University of Illinois Press, Urbana.
- Hennig W., 1969. Die Stammesgeschichte der Insekten. Senckenberg Buch 49, Verlag Waldemar Kramer, Frankfurt/M.
- Hennig W., 1971. Zur Situation der biologischen Systematik, in R. Siewing (dir.), Methoden der Phylogenetik, Erlanger Forschungen Reihe B 4, 7-15.
- Johnson L.A.S, 1970. Rainbow’s end : the quest for an optimal taxonomy. Systematic Zoology 19 : 203-239.
- Löther R., 1972. Die Beherrschung der Mannigfaltigkeit. Philosophische Grundlagen der Taxonomie. Gustav Fischer, Jena.
- Mayr E., 1974. Cladistic analysis or cladistic classification ? Zeitschrift für zoologische Systematik und Evolutionsforschung 12 : 95-128.
- Schlee D., 1971. Die Rekonstruktion der Phylogenese mit Hennigs Prinzip. Aufsätze und Reden der Senckenbergischen Naturforschenden Gesellschaft 20 : 1-62.
- Throckmorton L., 1965. Similarity versus relationship in Drosophila. Systematic Zoology 14 : 221-245.